5. Stav prírodného prostredia 10 rokov po rozdelení dunaja |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
|
vplyv zdrže na obsah rozpustených živín a na obsah živín viazaných na plaveniny, |
|
|
sezónne vplyvy na obsah živín, |
|
|
vplyv zvýšených prietokov a povodní na obsah živín. |
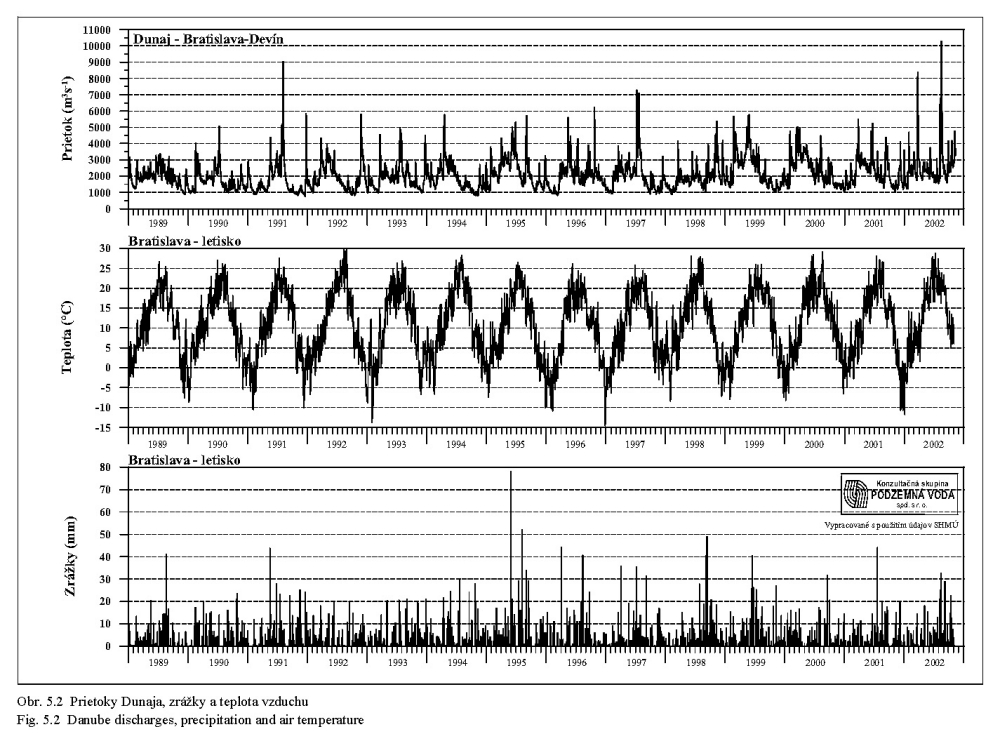
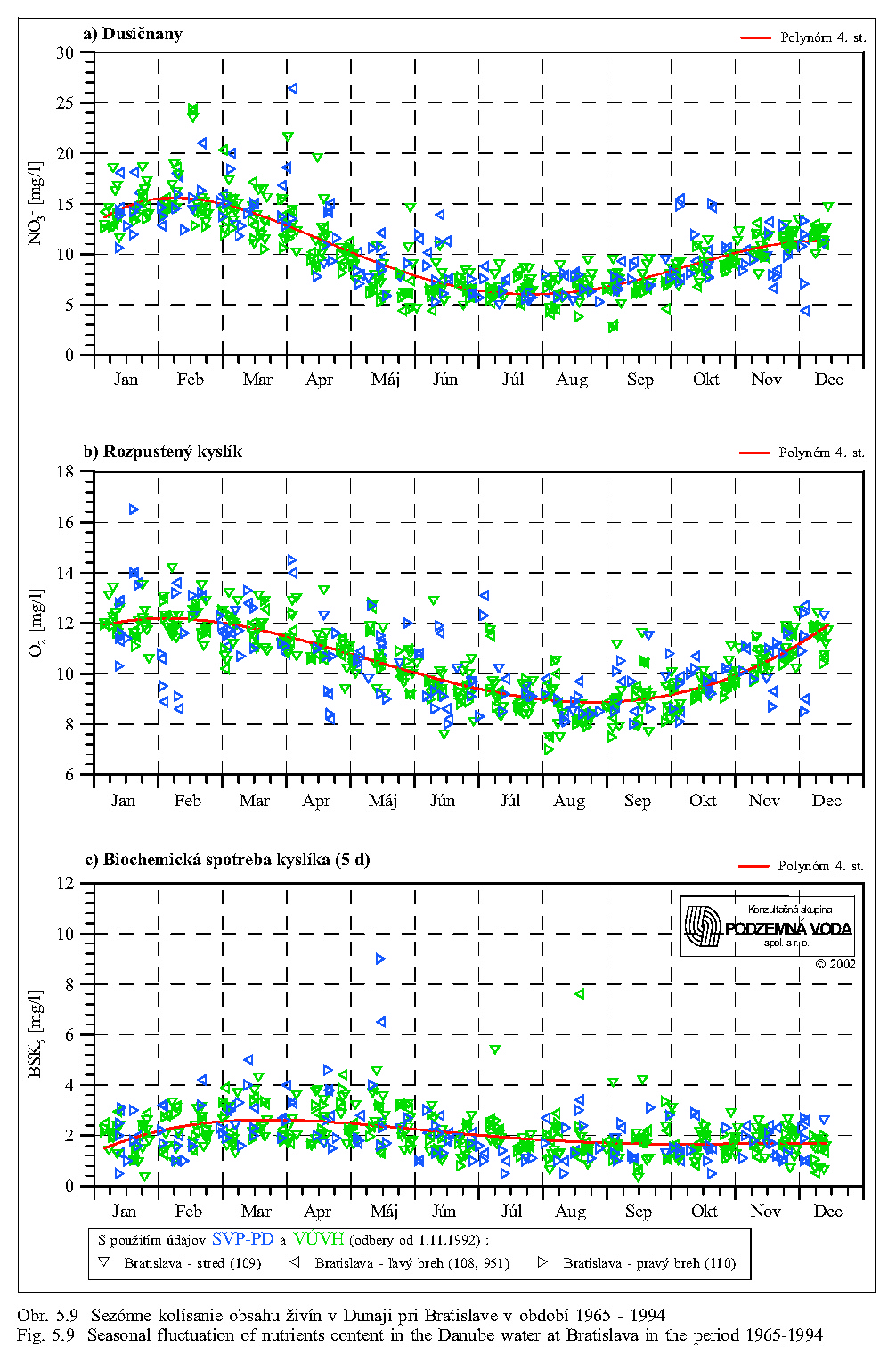
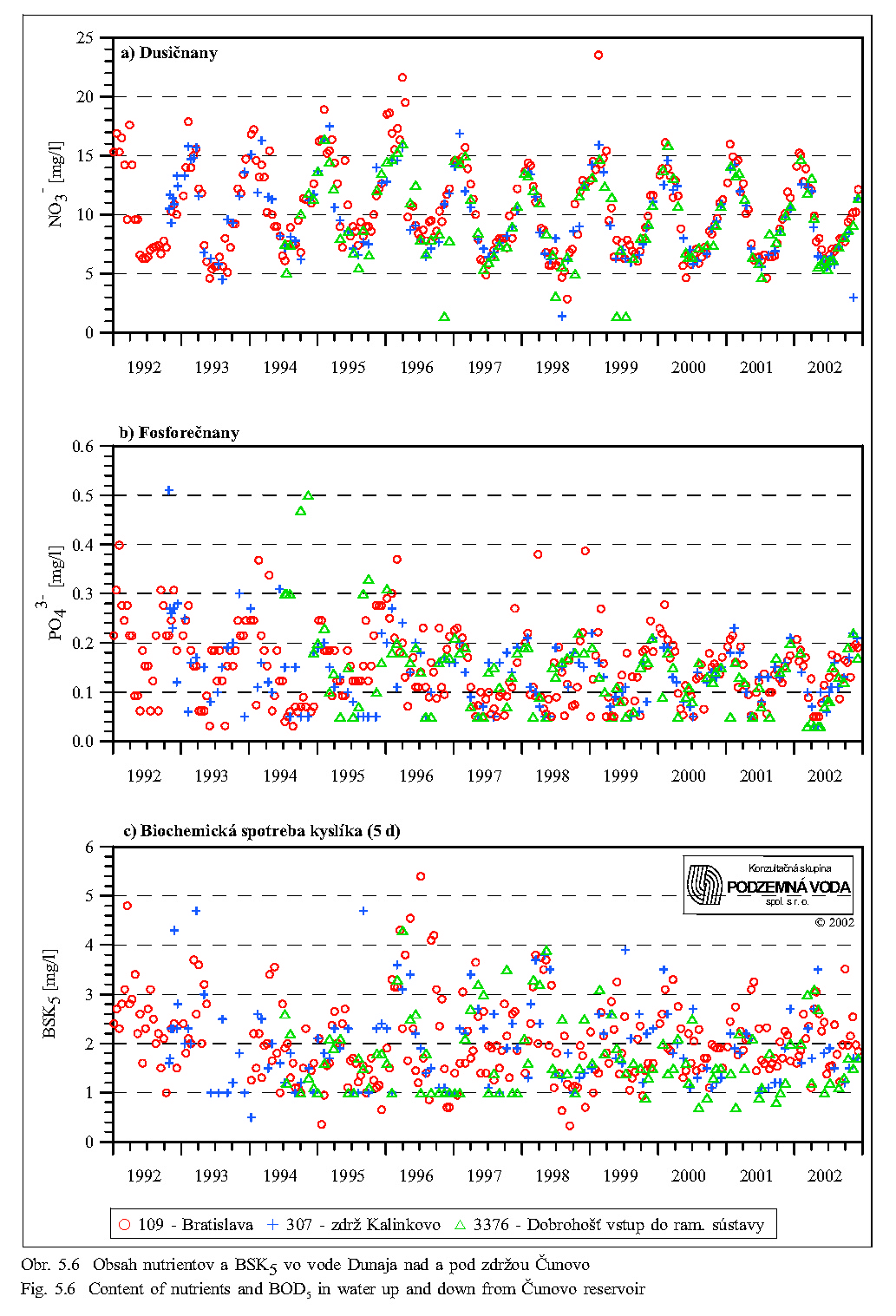
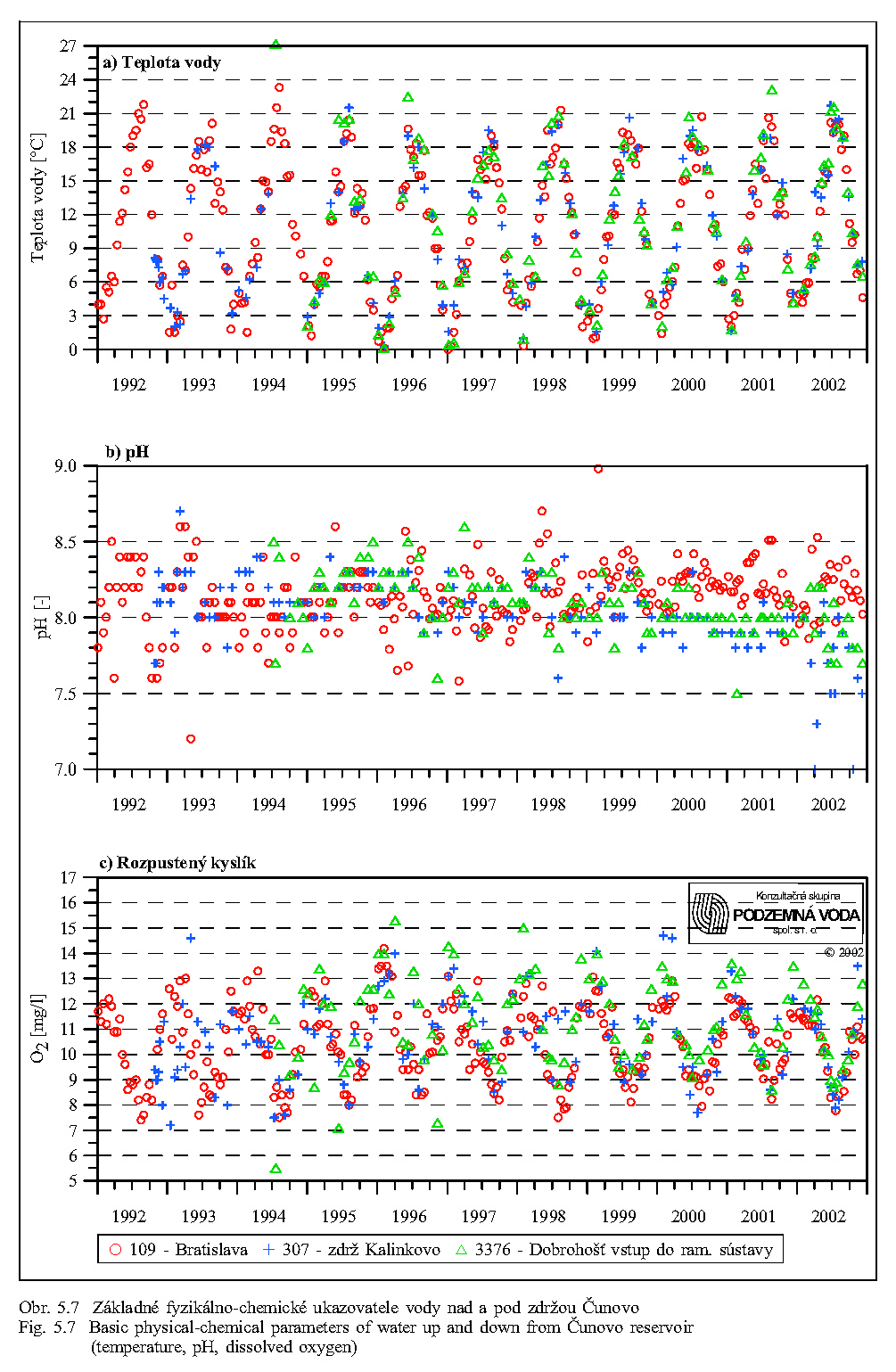
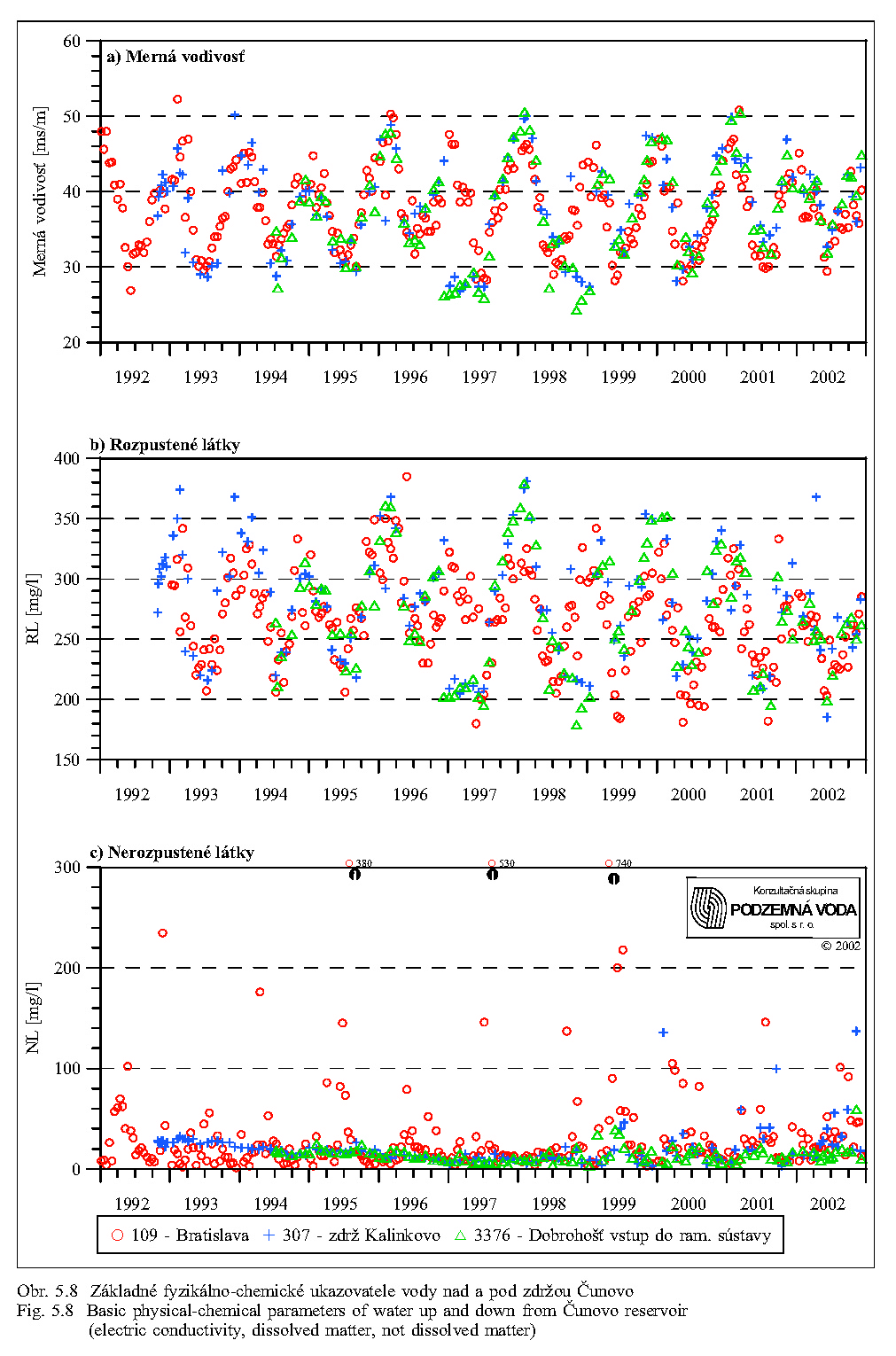
Séria grafov na Obr. 5.6, Obr. 5.7, Obr. 5.8 znázorňuje koncentrácie najdôležitejších živín v dunajskej vode v rokoch 1992 až 2000. Obsah živných látok rozpustených vo vode je charakterizovaný koncentráciami dusičnanov, fosforečnanov, hodnotou biochemickej spotreby kyslíka, obsahom rozpusteného kyslíka, elektrickou vodivosťou a celkovým obsahom rozpustených látok. Doplňujúcimi ukazovateľmi kvality vody sú teplota a pH vody.
{kind=link}
{kind=link}
{kind=link}
Obrázky dobre dokumentujú zmeny v obsahu rozpustných živín vplyvom prechodu vody cez zdrž Vodného diela Gabčíkovo. Pre porovnanie boli vybraté tri profily:
-
Dunaj v Bratislave (109), ktorý reprezentuje kvalitu vody pritekajúcej do zdrže,
-
zdrž pri Rusovciach/Kalinkove (307), ktorý reprezentuje kvalitu vody v hornej časti zdrže,
-
vstup do ramennej sústavy pri Dobrohošti (3376), ktorý reprezentuje kvalitu vody prepúšťanú do ramennej sústavy a zároveň vodu na začiatku prívodného kanála.
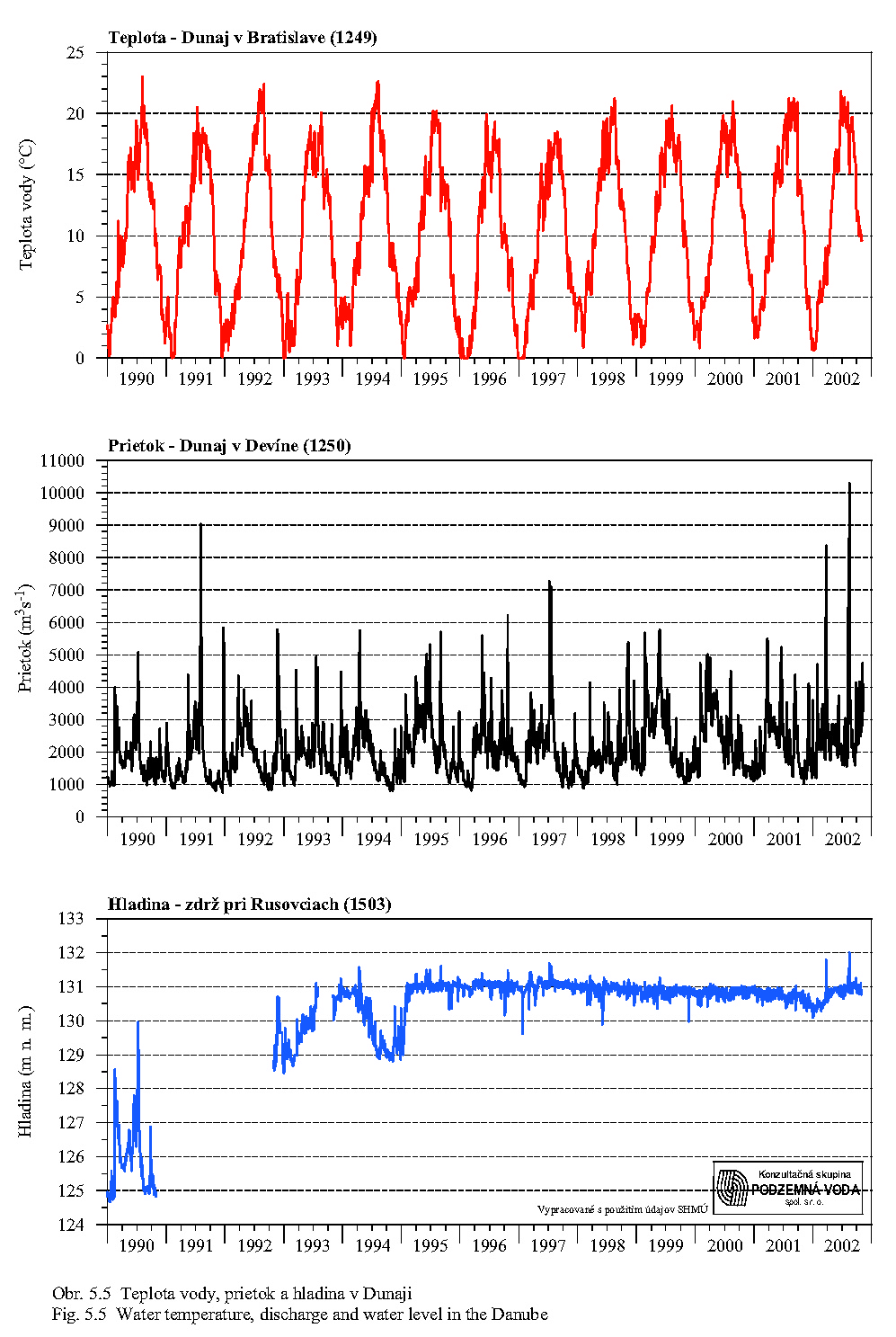
Z porovnania profilov nad a pod zdržou vidno, že prechod vody cez zdrž nespôsobuje signifikantné zmeny v obsahu rozpustných živín, ani zmenu pH a teploty vody.
Koncentrácia dusičnanov v tečúcej dunajskej vode sa mení behom roka v rozsahu asi 5-17 mg/l v závislosti na prietoku, obsahu organických látok a biologickej aktivite (porovnaj Obr. 5.6 a Obr. 5.9). Vyšší obsah dusičnanov sa vyskytuje mimo vegetačného obdobia v zime a počas jarného topenia snehu, najnižší je koncom leta a začiatkom jesene. Do biologických procesov na úrovni primárnych producentov vstupuje dusík najčastejšie v anorganickej forme, spravidla viazaný v dusičnanoch, u planktónnych rias skôr vo forme amoniaku (Lelák, Kubíček, 1991). Dusík sa nachádza v Dunaji obyčajne v dostatočnej miere a stáva sa nedostatkový len pri prebytku fosforu.
V porovnaní s inými biogénnymi prvkami má fosfor mimoriadny význam ako prvok často limitujúci produkčné procesy vo vodných ekosystémoch. Je to dané rozdielom medzi jeho pomerným zastúpením v živých organizmoch a jeho zdrojmi v prostredí (Lellák, Kubíček, 1991). Autotrofné organizmy zreteľne uprednostňujú fosfor vo forme rozpustených minerálnych fosforečnanov. K explózii sinicového vodného kvetu plne postačuje 10 μg fosforu v litri vody (Štěpánek, Červenka, 1974). Obsah fosforečnanov vykazuje podobné sezónne kolísanie ako dusičnany. Vo vode v oblasti nápustného objektu do ramennej sústavy sa v roku 1995 pohyboval okolo 0.2 mg/l (Obr. 5.6). Z toho vyplýva, že obsah fosforu vo vode Dunaja je väčšinou dostatočný a nie je inhibítorom biologických procesov. Do ramennej sústavy sa teda fosfor dostáva v dostatočnom množstve a za takmer každých podmienok.
Základná schéma kolobehu uhlíka vo vodných ekosystémoch vychádza z atmosféry; odtiaľ ho čerpajú producenti a odovzdávajú konzumentom; z oboch týchto trofických článkov postupuje uhlík k deštruentom. Uhlík prítomný vo forme anorganickej ako CO2 je využívaný pri asimilácii napríklad riasami. Biochemická spotreba kyslíka (BSK-5) charakterizuje nepriamo obsah organických látok, ktoré podliehajú biochemickému rozkladu pri aeróbnych podmienkach. Hodnota BSK-5 v dunajskej vode kolíše dlhodobo v rozsahu 1 až 3 mg O2/l, pričom je ovplyvňovaná viac prietokmi v Dunaji ako sezónnymi vplyvmi (Obr. 5.6, Obr. 5.9).
Kyslík je nevyhnutný pre rad dôležitých chemických i biochemických procesov a reakcií. Aktuálny obsah rozpusteného kyslíka v dunajskej vode pochádza zo vzduchu a z fotosyntetickej činnosti vodných rastlín. Difúzia kyslíka z ovzdušia do vody závisí najmä na teplote vody, najväčšia intenzita je v zime a na jar (Obr. 5.7). Priemerné kolísanie koncentrácií sa pohybuje v rozsahu 8 až 11 mg/l (Obr. 5.9). Kyslík spotrebovávajú pri dýchaní živočíchy i rastliny a jeho obsah vo vode kolíše významne aj v priebehu dňa. K poklesu koncentrácií rozpusteného kyslíka vplyvom respirácie dochádza počas noci, maximálne koncentrácie sa vyskytujú neskoro popoludní, kedy často dochádza k presýteniu vody kyslíkom.
Merná vodivosť vody (elektrická merná vodivosť) zodpovedá množstvu rozpustených látok disociovaných na ióny. Spolu s ukazovateľom celkového obsahu rozpustených látok predstavuje skupinový ukazovateľ, charakterizujúci obsah látok vo vode, bez určenia ich pôvodu a druhu. Obr. 5.8 ukazuje, že oba ukazovatele majú výrazne sezónny charakter (závisiaci hlavne od prietokov v Dunaji), maximálne hodnoty sa vyskytujú počas zimných mesiacov a skoro na jar, minimálne v lete.
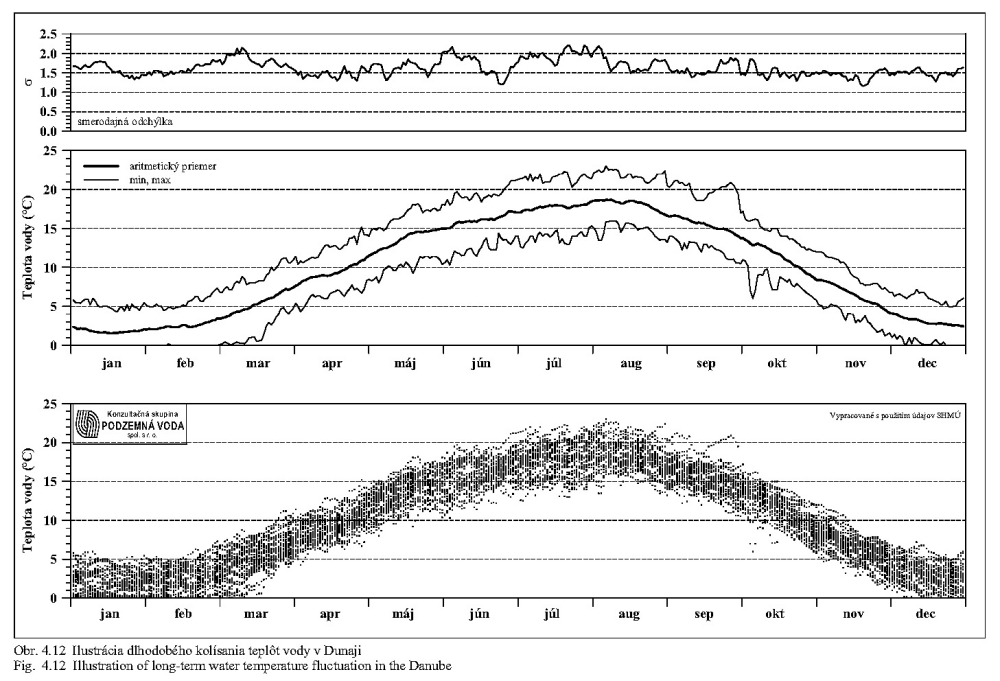
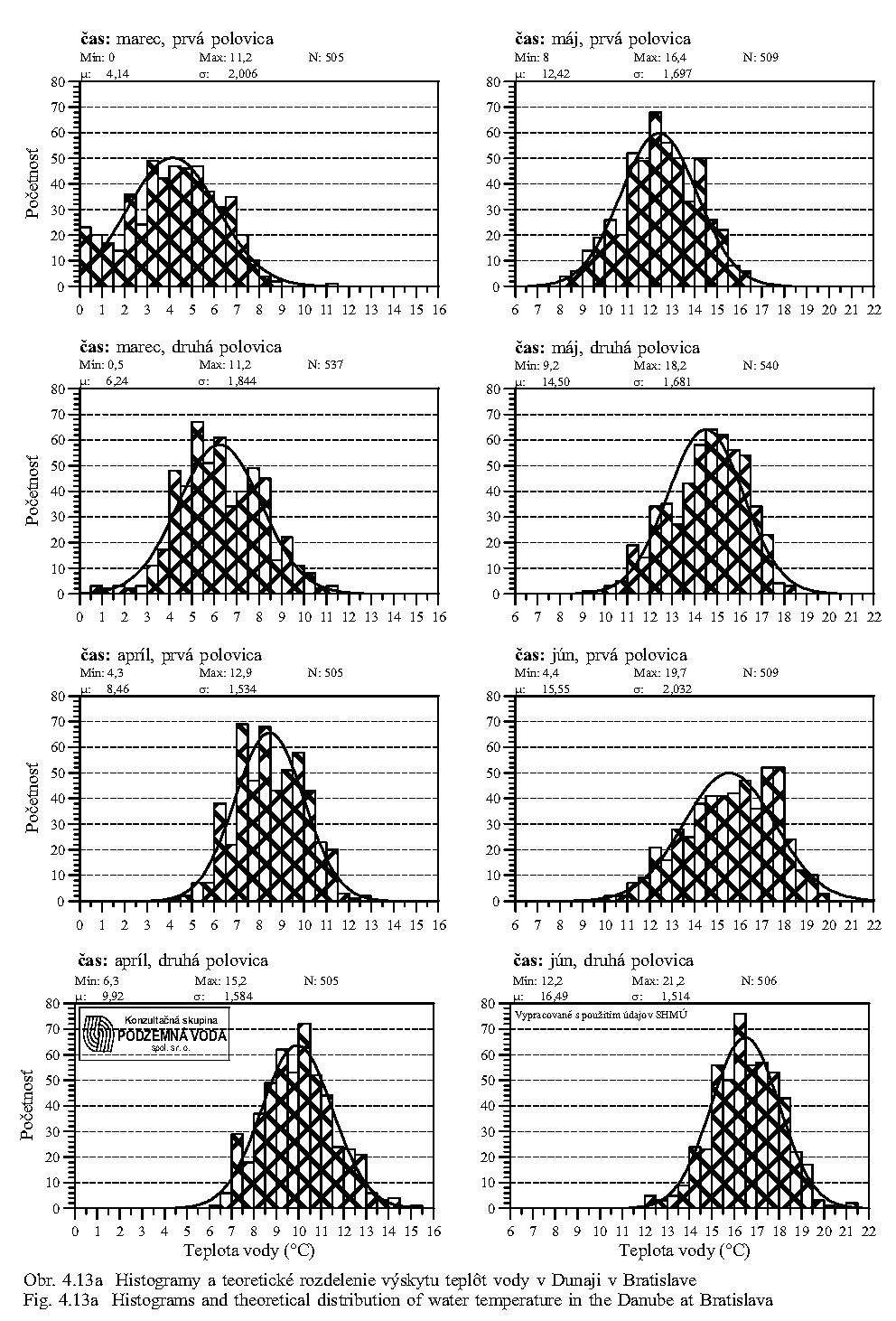
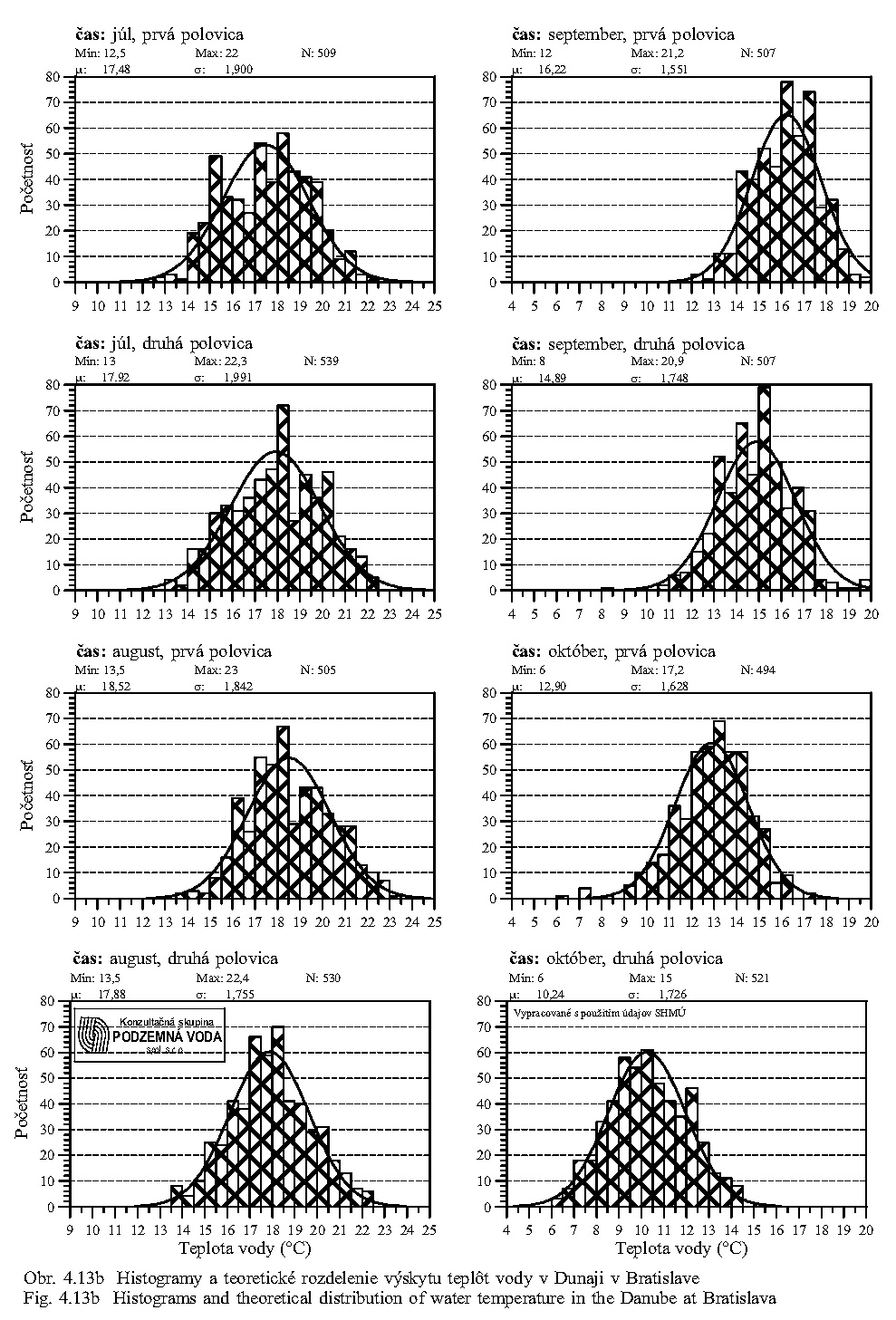
Teplota vody má všeobecný vplyv na rýchlosť metabolických procesov. Teplotné optimum pre väčšinu vodných organizmov je v rozpätí 10-30°C. Teplota vody v Dunaji kolíše v závislosti na sezóne v rozsahu od niekoľkých °C do 20 °C (Obr. 5.5). Ako vidno z Obr. 5.5, Obr. 5.7, Obr. 4.12, Obr. 4.13a, Obr. 4.13b, teplota vody v Dunaji dosahuje 10°C až v mesiacoch marec - apríl. V ramenách Dunaja sa voda prehrieva a pri malom prietoku až stojatej vode môže dosahovať pri slnečnom počasí podstatne vyššie hodnoty ako v hlavnom toku.
{kind=link}
{kind=link}
{kind=link}
Reakcia vody pH môže výrazne ovplyvniť oživenie vody. Niektoré druhy živočíchov znášajú veľké rozpätie pH vody, iné naopak len malé výkyvy pH v rozsahu desatín jednotky (Obr. 5.7). V prietočných ramenách pH vody odpovedá pH vode v Dunaji, v ramenách s malým prietokom a so stojatou vodou za určitých teplotných podmienok môže sa podstatne znížiť.
Koncentrácie ďalších rozpustených živín, napríklad draslíka, vápnika, horčíka a síranov majú podobný sezónny priebeh ako koncentrácie dusičnanov a rozpusteného kyslíka; maximálne hodnoty sa vyskytujú počas zimných mesiacov a skoro na jar, minimálne v lete.
Pri potrebe zvýšeného obsahu dôležitých rozpustených živín v dunajskej vode sa preto začiatok simulovanej záplavy na jar - v marci - javí ako najvhodnejší (pozri skupinové ukazovatele (elektrická) merná vodivosť (χ), rozpustené látky (RL), dusičnany, na Obr. 5.6, Obr. 5.8, Obr. 5.9).
Obsah živných látok transportovaných v suspendovanej forme ako plaveniny charakterizujeme celkovým obsahom nerozpustených látok (Obr. 5.8). Pevné látky usadené na dne alebo unášané vodou obsahujú anorganický i organický podiel, ktorý je potravou pre rôzne organizmy. V menej osídlených oblastiach prevláda anorganický materiál, v tokoch priemyslových a osídlených oblastí majú vyšší podiel látky organické, v poľnohospodársky využívaných oblastiach pôda. Dunaj je rieka s relatívne nízkym obsahom organických látok (Obr. 5.6, Obr. 5.9). Tieto látky sa počas povodní najviac zúčastňujú na zazemňovaní ramien s malými prietokmi a stagnujúcou vodou.
Ako vidno z obr. 5.8, obsah nerozpustených látok-plavenín je najvyšší v profile pri Bratislave. Vplyvom sedimentácie hrubozrnnejšej časti plavenín v zdrži sa postupne znižuje. V zdrži, v profile pri Rusovciach/Kalinkove, je menší a približne taký je aj v Dunaji v profile pri Dobrohošti a na odbernom objekte pri Dobrohošti. Porovnanie s Obr. 5.5 ukazuje, že obsah plavenín v profile pri Bratislave, a teda aj smerom po toku Dunaja, závisí významne od prietokov v Dunaji. Extrémne vysoké hodnoty nerozpustených látok sa vyskytujú v čase povodní.
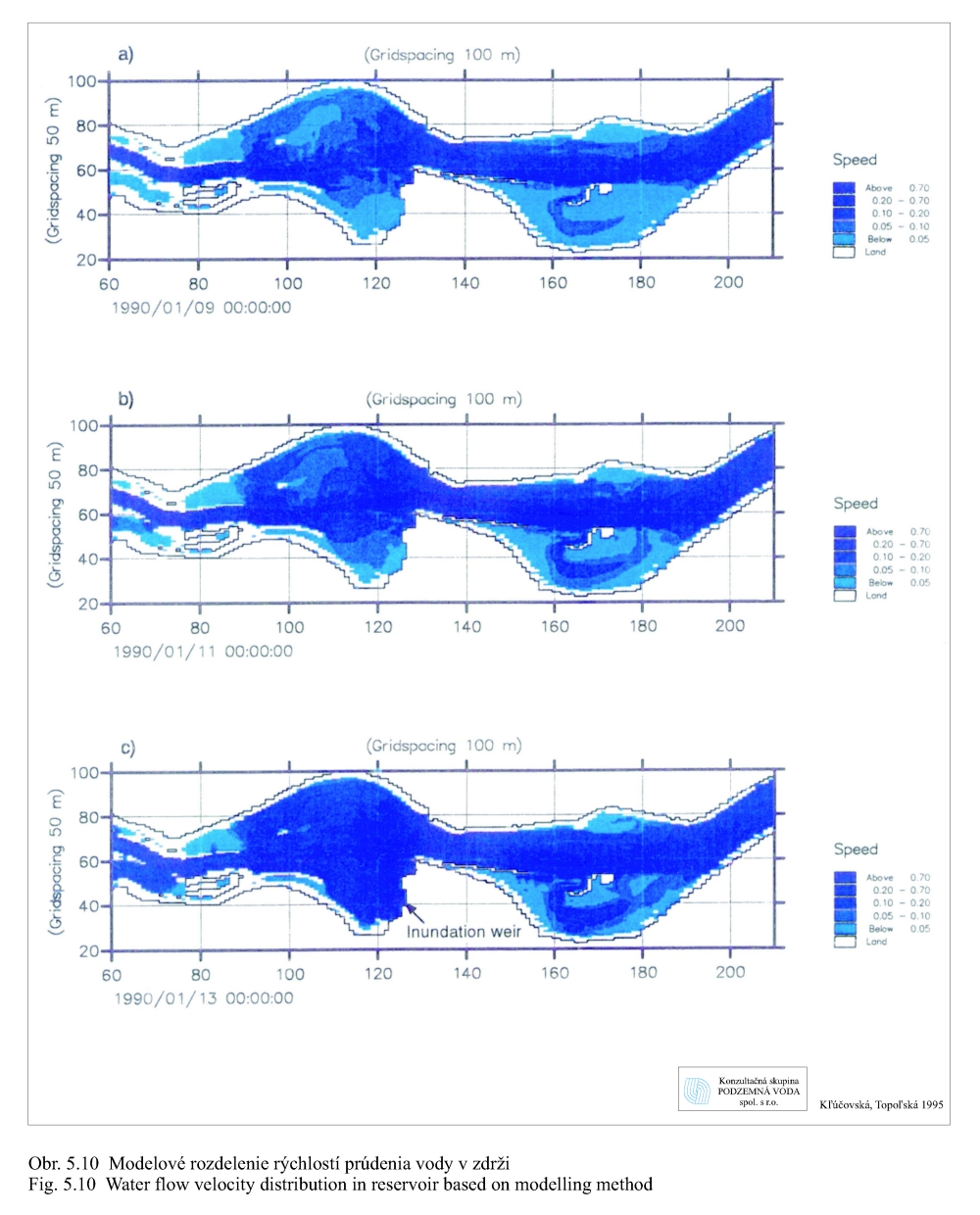
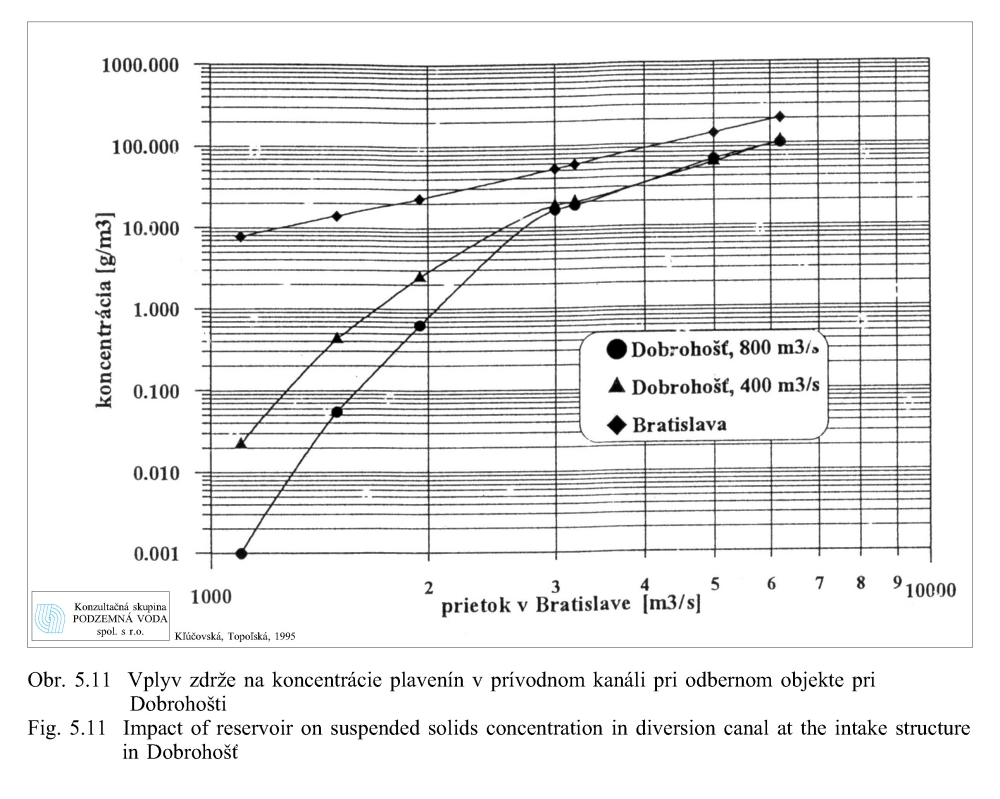
Transport plavenín závisí od prietokov a rýchlosti prúdenia vody. Obr. 5.10 zobrazuje modelované rozdelenie rýchlostí v zdrži pre rôzne prietoky v Bratislave: a) 1950 m3/s, b) 3.200 m3/s, c) 6.200 m3/s pri prepúšťaní 400 m3/s vody cez hať v Čunove do starého koryta Dunaja pri hladine 131.10 m n. m. (Kľúčovská, Topoľská 1995a, 1995b). Obrázky dokumentujú podmienky transportu plavenín (sedimentovanie, erózia/vznos) v zdrži pri rôznych prietokoch. Na obrázkoch vidno aj významný vplyv usmerňovacej hrádzky pri Šamoríne. Najvyššie rýchlosti prúdenia sa vyskytujú v miestach pôvodného koryta Dunaja a v plavebnej dráhe. Obr. 5.11 ukazuje modelovanú zmenu koncentrácie plavenín v závislosti na prietoku v Bratislave (Kľúčovská, Topoľská, 1995a, 1995b). Najvyššia koncentrácia plavenín je pri všetkých prietokoch v profile Bratislava. Pri priemernom prietoku 2.000 m3/s je koncentrácia plavenín v Bratislave 23 g v 1 m3 vody (23 mg/l) a koncentrácia plavenín pri nápustnom objekte do ramennej sústavy pri Dobrohošťi okolo 0.8 g/m3. Pri povodňovom prietoku 5.000 m3/s je koncentrácia plavenín v profile Bratislava okolo 120 g/m3 a pri Dobrohošťi sa zvýši na 65 g/m3. V čase povodní sa uvoľňujú pravdepodobne aj jemnozrnné sedimenty usadené na dne zdrže v miestach s vyššími rýchlosťami prúdenia vody (Obr. 5.10).
{kind=link}
{kind=link}
Z modelových riešení vyplýva, že záplavy ramennej sústavy vykonávané v čase prirodzených povodní cez nápustný objekt pri Dobrohošti zabezpečujú väčší prínos živín vo forme plavenín do ramennej sústavy, pretože v tom čase je obsah plavenín vo vode asi 80 krát vyšší ako pri priemernom prietoku v Dunaji. To by platilo ešte vo väčšej miere v prípade prepojenia ramien so starým korytom Dunaja.
Granulometria plavenín závisí od prietokov v Dunaji v Bratislave a od prietokov prevádzaných prívodným kanálom. Pri väčšom prietoku v Dunaji a väčšom prietoku cez derivačný kanál sa zvyšuje podiel hrubozrnnej frakcie plavenín v oblasti nápustného objektu pri Dobrohošti. Zrnitosť plavenín ovplyvňuje sorpčné procesy a priepustnosť dna cez plaveninové materiály po ich sedimentácii.
Z analýzy vyššie definovaných okruhov problémov možno na záver konštatovať, že:
|
Zdrž nespôsobuje signifikantné zmeny v obsahu rozpustených živín, ani zmenu pH a teploty vody prepúšťanej cez nápustný objekt pri Dobrohošti do ramennej sústavy. |
|
|
Koncentrácie rozpustných živín ako sú dusičnany, fosforečnany, kyslík, sírany, draslík, vápnik a horčík vykazujú výrazné sezónne kolísanie; maximálne hodnoty sa vyskytujú počas zimných mesiacov a skoro na jar, minimálne v lete. |
|
|
Zvýšené prietoky v Dunaji a povodne spôsobujú zvýšený transport plavenín z oblastí nad zdržou, nižšiu mieru sedimentácie plavenín v zdrži, a to i hrubozrnných, a čiastočne i uvoľňovanie najmä jemnozrnných plavenín z časti dna zdrže, čo znamená podstatný nárast v obsahu živín viazaných na plaveniny v porovnaní s priemernými vodnými stavmi. Prínos živín vo forme plavenín do ramennej sústavy cez nápustný objekt pri Dobrohošti je v čase povodne asi 80 krát vyšší ako pri priemernom prietoku v Dunaji. Povodňové plaveniny sa zúčastňujú veľkou mierou na zazemňovaní slepých ramien a ramien s obmedzeným prietokom. |
Termín simulovanej záplavy je možné voliť z hľadiska živín tak, aby do ramennej sústavy vtekala voda s určitým obsahom živín, či už v roztoku alebo viazaných na suspendované častice. Zároveň je možné voliť záplavu s väčším alebo menším obsahom suspendovaných častíc. Znamená to, že v období nízkych prietokov na Dunaji je možné väčším prepúšťaným množstvom vody preplachovať (čistiť) hlavné ramená ramennej sústavy. Z hľadiska zachovania prirodzených podmienok záplav je vhodné, aby termín simulovanej záplavy korešpondoval so zvýšenými prietokmi v Dunaji, najlepšie s prietokmi nad 4500 m3/s a s povodňovými stavmi na Dunaji
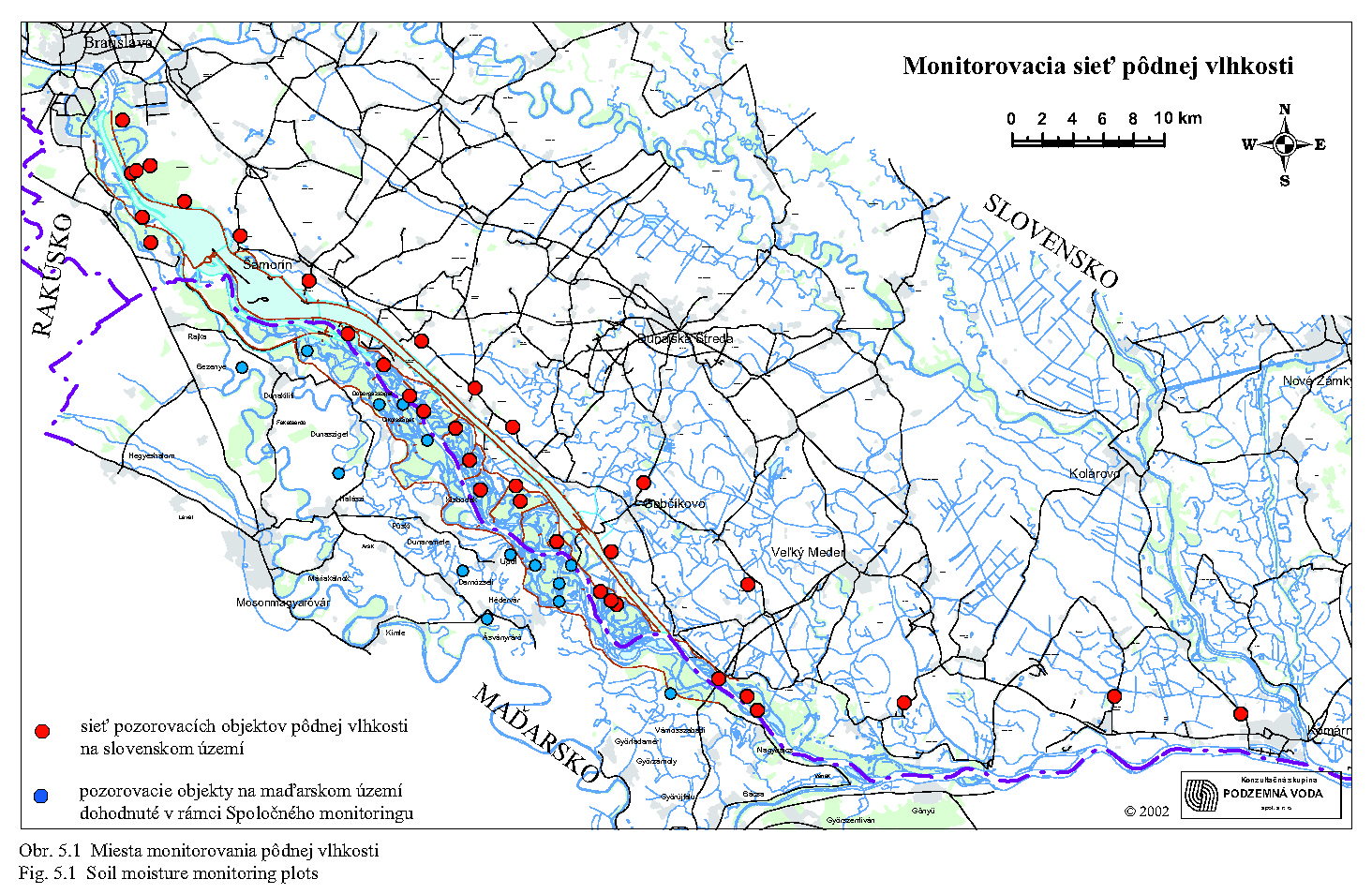
5.3 VÝSLEDKY MONITOROVANIA BIOTY
Monitoring rastlinstva (flóry a vegetácie) začal zisťovaním východiskového stavu na monitorovacích plochách (MP) založených v roku 1990 (Lisický et al., 1991). Z 24 založených monitorovacích plôch sa na inundačnom území nachádzali MP č. 6 (Dobrohošť), MP č. 7 (Žofín), MP č. 9 (Bodická brána), MP č. 10 (Kráľovská lúka), MP č. 14 (Istragov) a MP č. 15 (Erčéd), MP č. 17 (Diely), MP č. 18 (Sporná sihoť) a MP č. 23 (Čičov-Starý les). V podstate je to 9 monitorovacích plôch rozptýlených po rozlohe približne 3100 ha lužných lesov. Cieľom botanického monitoringu bolo získať základné aktualizované údaje o území z hľadiska makro-mezo a mikro-štruktúry vegetácie formou inventarizácie flóry a fytocenóz (Lisický et al., 1991). Po skonštatovaní východiskového stavu (pred prehradením Dunaja) v rokoch 1990-1992 boli vypustené MP č. 7, 17, 23 a väčšina spomínaných parametrov sa vyhodnocovala každoročne (Rovný et al., 1992, Cambel et al., 1993), i v rokoch po prehradení (Matečný et al., 1994, 1995, 1996, 1997, 1998, 1999, 2000, 2001). Súhrnné výsledky botanického monitoringu, zohľadňujúce i stratu olistenia a zmeny listovej plochy boli publikované v roku 1999 (Uherčíková et al., 1999). Vzhľadom na rozdielnu dynamiku vodného režimu v jednotlivých rokoch, ale aj zrážkových pomerov, sa výsledky vyznačujú značnou fluktuáciou počtu rastlinných populácií. Táto skutočnosť však nie je negatívnym dôsledkom výstavby VD, ale zákonitým javom inundačných území. Nepriamym monitoringom flóry (len na monitorovacích plochách a nie na celom území) bolo zistených v celom dotknutom území 760 cievnatých rastlinných druhov. Tento počet však zahrňuje floristický inventár i z takých monitorovacích plôch, ktoré sú situované mimo inundačného územia (Podunajské Biskupice, Rusovce, Čičov a i.). Žiaľ, od založenia monitorovacích plôch v roku 1990 sa celkový floristický inventár nevyhodnocoval. Zo štrukturálnych zmien vegetácie na monitorovacích plochách inundácie sa za sledované obdobie (1990-1995) urobili zovšeobecnenia (Uherčíková 2001), ktoré možno zhrnúť nasledovne:
|
zvýšené uplatňovanie sa neofytných rastlín s tendenciou ich udomácnenia, |
|
|
výrazné uplatňovanie nitrátofilných rastlín, |
|
|
absencia výrazne hydrofilných populácií, |
|
|
ochudobnenie druhového spektra väčšiny lesných spoločenstiev o 4 - 6 druhov. |
Z botanického monitoringu uplatňovaného cez odbornú skupinu „Biota“ za roky 1990-1996 vyplývajú závery vo forme odporúčaní akými sú napr.; zvýšiť hladinu vody v starom koryte Dunaja, doriešiť zásobovanie vodou v úseku Gabčíkovo-Prístav a sútok Dunaja s odpadovým kanálom a vzájomne poprepájať ramenný systém, nie však so starým korytom. Letné záplavy sa odporúčajú realizovať každý rok v mesiaci júl a august, v počte aspoň 33 dní, za účelom zvýšenia erozívno-akumulatívnej činnosti v rámci inundácie.
Z mapovania reálnej vegetácie inundácie (Šomšák et al., 2001, 2002) bolo urobené porovnanie rozlohy jednotlivých typov vŕbovo-topoľového lesa v roku 1959 a 2001. Výsledky ukazujú takmer zhodnú rozlohu. Tento fakt spolu s porovnaním oscilácie hladín podzemných vôd v rokoch 1951-1953 (Jurko 1958) a v súčasnosti (Mucha et al. 2001) navádzajú k presvedčeniu, že súčasný hydropedologický režim zavodňovania ramennej sústav inundácie je z hľadiska terestrickej vegetácie takmer optimálny (Šomšák 2001). Chýba tu však erozívno-akumulatívna činnosť ako podmienka prírodnej obnovy vŕbovo-topoľových lesov. Túto však možno dosiahnuť (i keď v menšej miere) simulovanými záplavami v jarnom období.
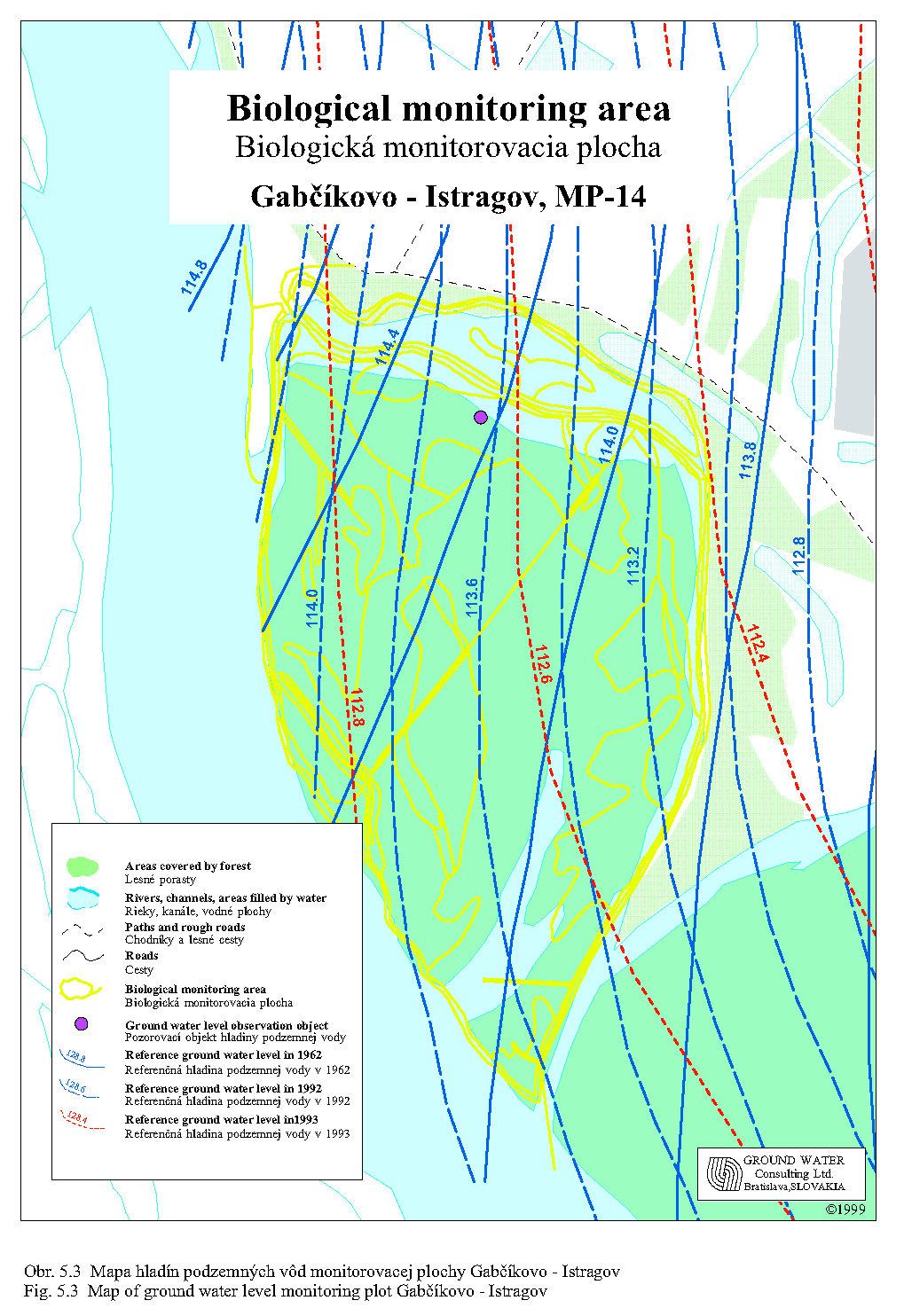
Rozloženie trvalých monitorovacích plôch (TMP) lesníckeho výskumu odráža reprezentatívnejšie situáciu hydrologického režimu inundácie. Monitorovacích plôch je 10, pričom 8 z nich monitoruje situáciu medzi obcou Dobrohošť a vyústením Bačianského ramena do Dunaja a dve sledujú vývoj lesných porastov medzi Gabčíkovským prístavom a sútokom odpadového kanála so starým Dunajom. TMP v hornom úseku inundácie sú lokalizované tak, aby v každej sekcii medzi dvoma prehrádzkovými líniami bola aspoň jedna a aby v celom sledovanom úseku reprezentovali klonovú skladbu drevín a vekovú štruktúru, v čase ich založenia (Neštický, Varga 2001). V roku 1997 bolo z hľadiska prírastkov drevnej hmoty a zdravotného stavu lesa vyhodnocovaných ešte ďalších 11 pokusných plôch, nezahrnutých do oficiálne evidovaného lesníckeho monitoringu. Údaje z monitorovania za roky 1993-1996 boli zhrnuté do súbornej záverečnej správy (Varga et al., 1997). Autori tejto správy rozdelili inundačné územie z hľadiska dynamiky hladín podzemných vôd vo vzťahu k lesným porastom na 5 skupín:
Územie od nápustného objektu po líniu „B“. Hladina podzemných vôd je v tejto skupine porastov pre zdravý vývoj a očakávanú produkciu drevnej hmoty nevyhovujúca. Optimalizáciu možno, podľa autorov, dosiahnuť vybudovaním dnových prehrádzok v starom koryte Dunaja.
Príbrežný pás popri starom koryte Dunaja v celom úseku derivácie. Pôdy sú tu drénované starým korytom. Lesohospodárska optimalizácia je možná len zvýšením hladiny v starom koryte Dunaja, alebo prestavbou drevinového zloženia existujúcich porastov.
Územie od línie „B“ po vyústenie Bačianského ramena do starého koryta. Podľa autorov (Varga et al. 1997, Neštický, Varga 2001) je to územie, kde dynamika hladiny podzemnej vody je pre vývoj zdravotného stavu lesa a produkciu drevnej hmoty vyhovujúca.
Územie od vyústenia Bačianského ramena do starého koryta Dunaja po ústie odpadového kanála do Dunaja (Istragov). Dynamika hladín podzemnej vody vo vzťahu k lesným porastom je veľmi variabilná, závisiaca na súčinnosti prietokov v odpadovom kanáli a v starom koryte Dunaja. Pravidelne sa opakujúce zníženie hladiny podzemnej vody vo vegetačnom období bude potrebné eliminovať osobitnou dotáciou vody do celého ramenného systému Istragova.
Porasty v blízkosti ľavobrežnej hrádze prívodového a odpadového kanála. Vzhľadom na to, že táto časť nebola vyhodnocovaná monitoringom „Biota“ (Uherčíková et al., 1999), hodnoty hladiny podzemnej vody uvádzané na základe lesníckych pozorovaní (Varga et al., 1997) budú mať význam pre lesopestovateľskú prax (obnova lesa a rekonštrukcia lesných porastov podľa nárokov na vlhkosť pôdy, a i.).
Výsledky takmer všetkých lesníckych pozorovaní z inundačného územia zdôrazňujú nevyhnutnosť simulovaných záplav. Tieto sú významné z hľadiska dostatočného nasýtenia pôdnych profilov aj tých stanovíšť, kde je hladina podzemných vôd dostatočne vysoko, a to nielen z hľadiska vlahy, ale i ako zdroja prísunu živín (Neštický et al., 1996, Varga et al., 1997, Neštický, Varga 2001). Nevyhnutnosť simulovaných záplav ako hlavnej podmienky pre prírodnú obnovu vŕbovo-topoľových lesov zdôrazňujú aj iní autori (Šomšák 1998, 1999). Dlhodobým sledovaním prírodnej obnovy vŕb a topoľov (Šomšák, 1998, 1999, Pišút, Uherčíková 1993) bolo zistené, že obnova pôvodných drevín (Salix alba, Populus nigra, Populus alba) zo semena prebieha len na obnažených sedimentoch, alebo na záplavami oderodovanom fluvizemnom podklade. Obnova zo semien prebieha vždy spontánne, so znakmi viacročných fluktuácií. Obnova pod materským porastom zo semena týchto drevín bola a aj v súčasnosti je veľmi zriedkavá.
Z ekologických syntéz vzťahujúcich sa na rastlinstvo (flóru a vegetáciu) má rozhodujúci význam gradientová analýza spracovaná podľa programového balíka CANOCO. Pre záujmové územie ho aplikovala Uherčíková (1995). V analýze išlo o vyjadrenie vzťahu floristického zloženia lesných fytocenóz k známym veličinám prostredia. Za premenné prostredie tu autorka považovala namerané hodnoty 9 faktorov (dusičnanový dusík, celkový dusík, pH, H2O, CaCO3, humus, zrnitosť rizosféry, vek porastu a dve referenčné hladiny podzemnej vody z roku 1962 a 1992). Najsilnejšie korelovaným faktorom bola referenčná hladina podzemných vôd. Potvrdila sa očakávaná a známa závislosť, že výška hladiny podzemnej vody je rozhodujúcim činiteľom distribúcie fytocenóz lužného lesa. Na základe výsledkov gradientovej analýzy treba upozorniť na narastanie druhového spektra nitrátofilných rastlinných populácií, postupne potláčajúcich prírodnú biodiverzitu rastlinstva. I keď za príčinu je považované uvoľňovanie zápoja stromových korún a zvýšeného prieniku svetla do bylinného podrastu podporujúce zrýchlenie rozkladu nahromadených organických zvyškov, nie je možné zabudnúť i na dôsledok absencie záplav a ch sprievodného erozívno-akumulatívneho procesu. Nežiadúcim zmenám by zabránilo opakovanie prirodzených i väčších simulovaných záplav.
Charakteristika fytodiverzity
Keďže v inundačnom území posudzovaného úseku Dunaja majú absolútnu plošnú prevahu vŕbovo-topoľové lesy (Salici-Populetum) bola diverzita posudzovaná len pre toto spoločenstvo. Vybraný bol typický vŕbovo-topoľový ekosystém (subasociácia Salici – Populetum typicum Jurko 1958). Pre porovnanie bolo vybraných z roku 1958 dvadsaťpäť zápisov (Jurko 1958) a 45 zápisov z toho istého spoločenstva z roku 1995 (Uherčíková 1995). Biodiverzita rastlinstva sa teda porovnávala po 35 rokoch (z toho tri roky po prehradení Dunaja).
Na porovnanie diverzity sa použil index škálovej diverzity (Dsc) a Hillov index diverzity (Mičuda 2002). Index škálovej diverzity preto, lebo je najvhodnejší pre porovnávanie vegetačných typov navzájom ako aj pre mapovanie diverzity v krajine. Index Dsc vychádza zo súčtu bodov priemerných hodnôt počtu druhov (xi) a priemernej pokryvnosti (yi) každej vrstvy. Hillov index diverzity sa odporúča v praxi nakoľko dáva široké a prehľadné spektrum hodnôt. Maximálna hodnota je 100 a minimálna je 1. Hillov index diverzity vyjadruje kvadratický podiel koeficientu významnosti každého druhu.
Výsledky porovnania:
|
Zníženie škálovej diverzity o stupeň (zo stupňa 7 – vysoká v roku 1958 (so zápisov Jurko 1958), na stupeň 6 – stredná až vysoká škálová diverzita v roku 1995 (so zápisov Uherčíková 1995). |
|
|
Zníženie Hillovho indexu diverzity z hodnoty 42,1 v roku 1958 (so zápisov Jurko 1958) na 39,1 v roku 1995 (so zápisov Uherčíková 1995). |
|
|
Zníženie priemernej pokryvnosti v roku 1995 (so zápisov Uherčíková 1995) oproti roku 1959 (so zápisov Jurko 1959) vo všetkých etážach. |
|
|
Malý nárast priemerného počtu druhov v stromovej a krovinnej etáži a pokles v bylinnej vrstve v roku 1995 oproti roku 1958. |
Zníženie škálovej diverzity o 1 stupeň a zníženie Hillovho indexu diverzity o 3 body nedovoľuje urobiť závery o vážnejších zmenách. S prihliadnutím k výsledkom gradientovej analýzy, ktorá potvrdila závislosť týchto ekosystémov na dynamike hladín podzemných vôd (Uherčíková 1995) možno však konštatovať, že simulovaný vodný režim inundačného územia je v hrubých rysoch pre väčšiu časť lesov vyhovujúci. Znovu podčiarkujeme, že to neplatí pre drenážovanú pobrežnú časť okolo starého koryta Dunaja, ktorý musí byť riešený osobitne.
Súčasný stav a zmeny fauny a živočíšnych taxocenóz, ktoré sa odohrali na území ovplyvnenom výstavbou a prevádzkovaním VD Gabčíkovo, posudzujeme na základe výsledkov monitoringu bioty v rokoch 1990-1997 (Lisický et al., 1991, Rovný et al., 1992, Cambel et al., 1993, Matečný et al., 1994 až 2001). Východiskový stav je opísaný v prvých troch z nich. Monitoring sa robil, rovnako ako pre flóru, v oblasti derivácie najskôr na 9 a od roku 1991 na 5 monitorovacích plochách. Plochy majú rozličný hladinový režim povrchových i podzemných vôd. V rámci monitoringu terestrickej fauny bol sledovaný celý rad taxocenóz pôdy, epigeonu a nadzemných etáží (Mollusca, Oniscidea, Acari, Chilopoda, Collembola, Heteroptera, Coleoptera, Neuroptera, Mecoptera, Lepidoptera, Hymenoptera, Amphibia, Aves, Mammalia a i.), ktoré pomerne výrazne odrážajú zmeny a stav v spoločenstvách terestrických a semiakvikolných živočíchov.
Zmeny taxocenóz treba posudzovať z niekoľkých hľadísk. Prvým z nich je zmena druhového bohatstva spoločenstiev, avšak samotná zmena počtu druhov nepredstavuje všeobecne a v lužných lesoch zvlášť, príliš významné indikačné kritérium, pretože v procese degradatívnych zmien spoločenstva sú miznúce druhy charakteristické pre prirodzené pomery nahrádzané približne rovnakým, resp. aj vyšším, počtom xenocénnych druhov. Preto sa celkový počet druhov významne nemení, resp. môže stúpnuť. Tento jav je niekedy nesprávne interpretovaný (aj v prípade zmien vo vnútrozemskej delte Dunaja), ako zlepšenie ekologickej situácie v dôsledku antropických zásahov. Pre správnu interpretáciu zmien počtu druhov je nevyhnuté zohľadnenie ekologických vlastností jednotlivých druhov. Zvlášť významný je pomer zastúpenia druhov s rozličnými vlhkostnými nárokmi, ktorý však závisí na stupni poznania autekológie druhov. Z hľadiska potrieb bioindikácie zmien živočíšnych spoločenstiev v lužných lesoch ide o najvýznamnejšie kritériá s vysokou výpovednou hodnotou. Druhým významným kritériom je pomer druhov vyžadujúcich trvalé zatienenie drevinnou vegetáciou k druhom indiferentným alebo vyžadujúcim nelesné ekosystémy bez krovinovej a korunovej etáže. V lužných lesoch je to kritérium reflektujúce druhotné, ale synergicky pôsobiace zmeny, vyvolané primárne zmenami vlhkosti (presychanie porastov, úbytok vlhkomilných drevín a z toho vyplývajúce preriedenie zapojenia korún), vplyv abiotických činiteľov (vývraty spôsobené vetrom), alebo ľudskými zásahmi (pestovateľské lesohospodárske zásahy - prerezávky, výberová ťažba).
Súčasné zloženie taxocenóz je výsledkom zmien, ktoré sa na tomto území odohrali počas výstavby VD Gabčíkovo a po jeho sprevádzkovaní. Drénový efekt pôvodného koryta spôsobil zhoršenie stanovištných podmienok mäkkého lužného lesa. Významné z hľadiska fauny je, že presychajú vrbiny pobrežného pásu a vysychajú v minulosti vodnaté depresie a ramená, ktoré nekomunikujú s dotovanou ramennou sústavou. V páse pozdĺž starého koryta Dunaja, ktorého šírka sa mení podľa konfigurácie hlavných ramien (80-250 m), hladina podzemných vôd nie je v kontakte s kapilárnou obrubou rizosféry drevín. Dochádza k predčasnému opadu listov a poloprirodzené i hospodárske porasty odumierajú (Pišút 1995), dochádza k aridizácii územia, do ktorého v najväčšej miere prenikajú nepôvodné faunistické prvky (Jedlička et al., 1999).
Zo živočíšnych taxocenóz na tieto zmeny reagujú najmä edafické a epigeické taxocenózy, ktoré sú viac viazané na pôdne podmienky, najmä vlhkostné. Taxocenózy nadzemných etáží sú v istej miere viac závislé na klimatických podmienkach (vrátane mikroklímy) a vegetácii (Jedlička et al., 1999).
Územie predstavuje v istom zmysle ostrov, protiprúdne izolovaný od zázemia vodným dielom. Významnejšou skutočnosťou však je, že terestrické zoocenózy lužných lesov v úseku Bratislava – Dobrohošť sú značne degradované, ochudobnené. Napr. v malakocenóze aj na najvlhších stanovištiach spoločenstiev Salici-Populetum chýbajú i menej náročné vlhkomilné druhy, alebo sa vyskytujú len vo veľmi nízkej početnosti, ktoré sa východnejšie (poprúdne) vyskytujú v početných populáciách (Ševčíková 1997, Čejka 1999, Jedlička et al., 1999). Vo väčšine lužných lesov pri Bratislave sú nahradené hlavne lesnými mezohygrofilnými a niektorými hygrofilnými druhmi. V zachovalejších typoch vlhších porastov je to predovšetkým Clausilia pumila a Semilimax semilimax (zriedkavejšie aj Vitrea crystallina), v suchších typoch zasa druhy ilýrskeho lesného regiotypu (najmä Aegopinella nitens, ďalej Petasina unidentata, Monachoides incarnatus, Cochlodina laminata) a niektoré druhy regiotypu prevažne lesných starousadlíkov (Alinda biplicata, ale aj Fruticicola fruticum a na vlhších miestach tiež Arianta arbustorum). Dôvodom takejto štruktúry taxocenóz je dlhodobý pokles hladiny podzemnej vody v súvislosti so zahlbovaním koryta rieky. V dôsledku toho došlo k vymiznutiu polyhygrofilných druhov a prípadné zvyškové populácie už neskôr neboli schopné obnovy. Potenciálne imigračné zdroje predstavujú dunajské náplavy. Imigranti tu však zrejme nenachádzajú vhodné podmienky, prípadne sú naplavené populácie tak málo početné, že nie sú schopné ďalšieho samostatného vývoja (Čejka a Falťan 2001). Podobná situácia je aj v ostatných skromných zvyškoch mäkkých luhov pod Bratislavou, a to aj v tej oblasti, ktorá bola pozitívne ovplyvnená vzdutím hladiny podzemnej vody prostredníctvom čunovskej zdrže. Toto územie nemôže slúžiť ako prípadné refúgium a zdroj migrácie hygrofilných ekoelementov do oblasti derivácie. Oblasť derivácie je podobným spôsobom izolovaná aj z ostatných strán kultúrnou stepou (poľnohospodárskou krajinou) a len pod vyústením odpadového kanála nadväzuje na ekologicky podobné územie malého rozsahu. Vzhľadom na známy vzťah druhového bohatstva (biodiverzity) a plochy a teóriu ostrovov je pre zachovanie biodiverzity potrebné v plnom rozsahu zachovať rozsah spoločenstiev vnútrozemskej delty.
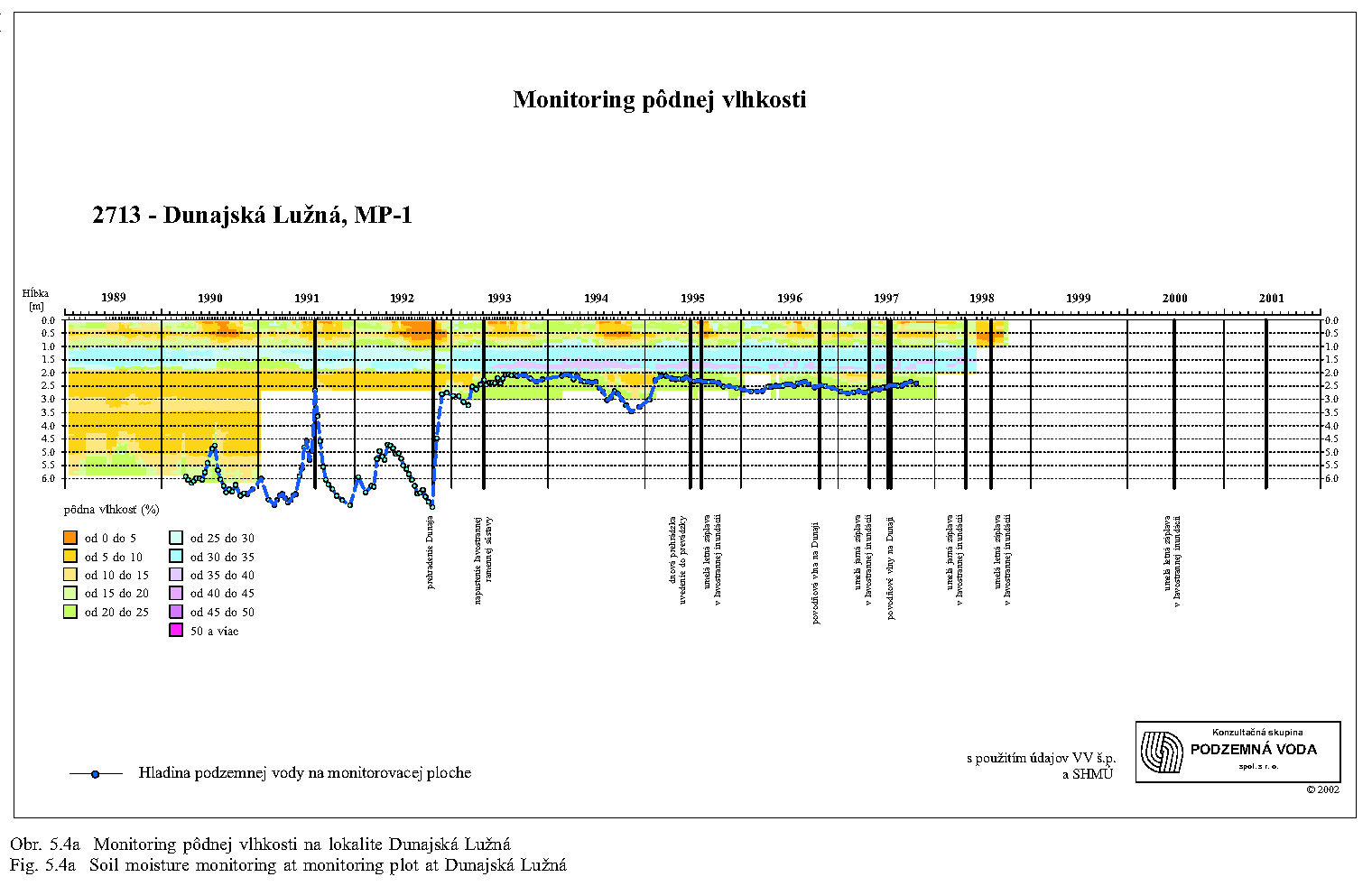
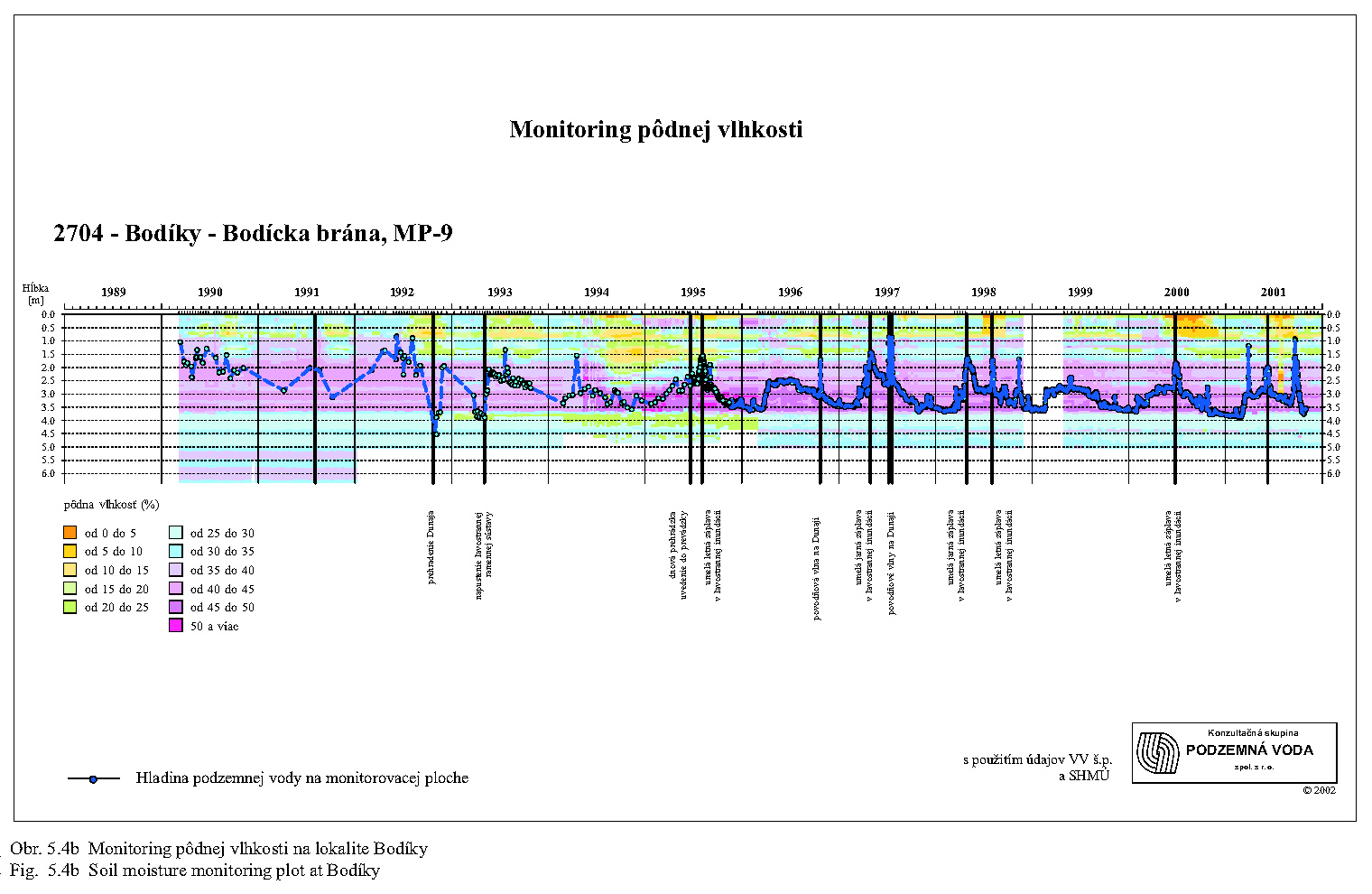
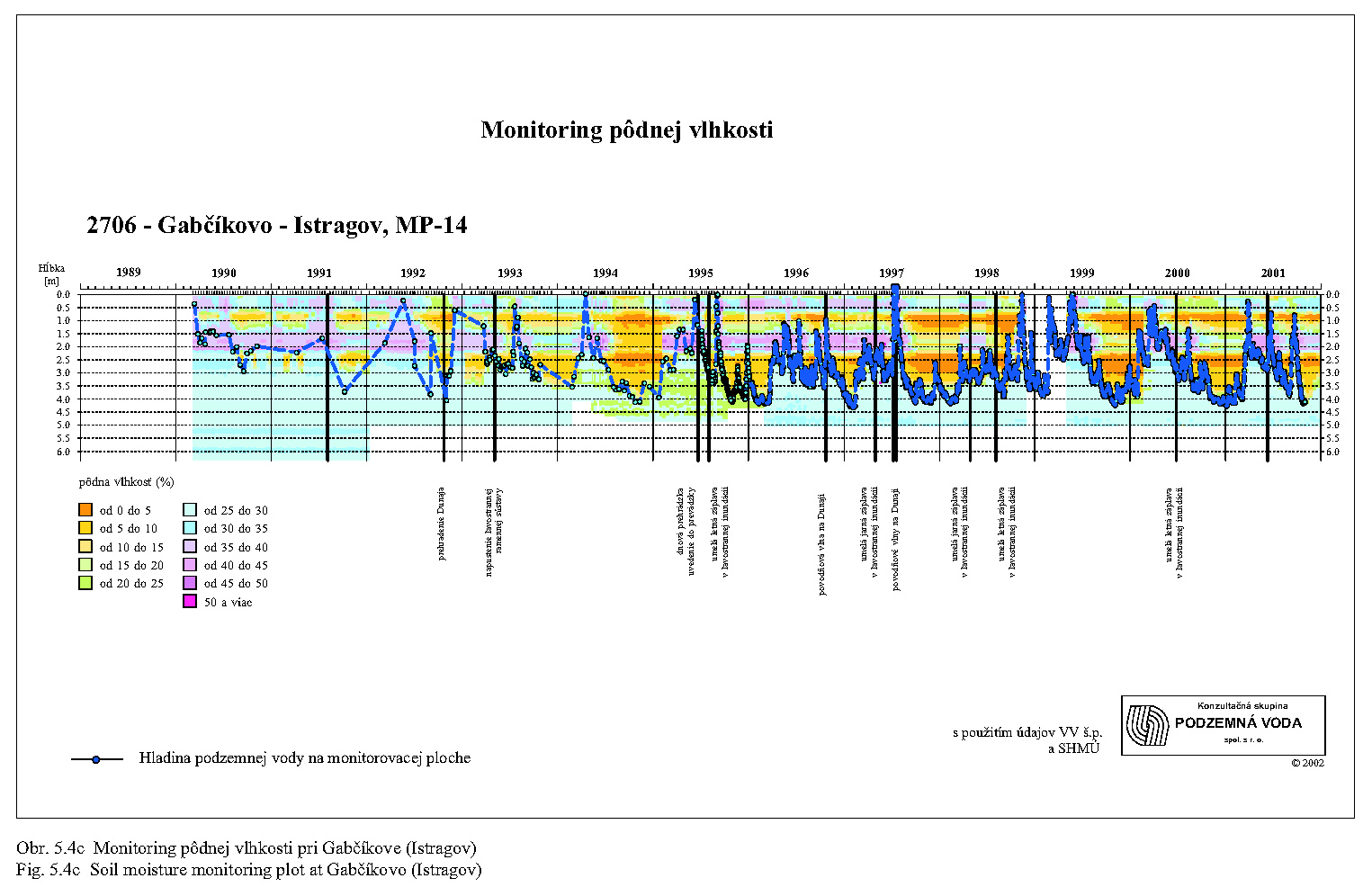
Malakocenózy aj v dôsledku nízkej vagility reagujú pomerne citlivo na zmenu podmienok. Najväčšie zmeny v štruktúre malakocenóz v zmysle ubúdania hygrofilných druhov pozorujeme po prehradení Dunaja najmä v oblasti Dobrohošťa (monitorovacia plocha Dunajské kriviny), Bodíkov (Bodícka brána) a Gabčíkova (Istragov). Pôvodné malakocenózy mäkkého lužného lesa s prevahou výrazne hygrofilných druhov sa menia na mezohygrofilné cenózy charakteristické skôr pre prechodný až tvrdý lužný les, najmä pri Dobrohošti. Na prelome 80. a 90. rokov tu existovala malakocenóza typická pre stredne vlhké až vlhké varianty mäkkého luhu s dominantnými polyhygrofilnými druhmi Carychium minimum, Zonitoides nitidus a Succinea putris. Podľa rozboru tanatocenóz z pôdnych vzoriek tu v minulosti žil aj mokraďový druh Vertigo antivertigo, ktorý nebol z Podunajska hlásený už zo dve desaťročia. Po prehradení Dunaja na monitorovacej ploche pri Dobrohošti takéto druhy populácie postupne vymizli.
V oblasti pri Bodíkoch je situácia o čosi priaznivejšia. Po prehradení Dunaja bývajú stanovištia sporadicky zaplavované, reštituovanie podielu vlhkomilných druhov (C. minimum, Z. nitidus, V. crystallina) na úroveň pred prehradením sme však zatiaľ nezistili. Krátkodobé simulované záplavy ovplyvňujú vlhkostné pomery len na krátky čas, vzhľadom na dominantný účinok drénu starého koryta a piesčitý substrát. Musíme preto konštatovať, že štruktúra malakocenóz nie je manipuláciou s vodou v inundačnom území pozitívne ovplyvnená. V pôvodných vlhkomilných taxocenózach, ktoré v tejto oblasti existovali pred prehradením Dunaja, sa na úkor polyhygrofilných druhov zvyšuje postupne podiel mezohygrofilných a euryekných druhov (A. nitens, M. incarnatus, Alinda biplicata, Punctum pygmaeum). Monitorované taxocenózy na lokalite Istragov pri Gabčíkove patrili do roku 1992 k výrazne vlhkomilným. Monitorovacia plocha bola jediná, kde sa pred prehradením vyskytoval vzácny mokraďový druh Euconulus alderi. Po prehradení Dunaja a znížení prietoku v jeho pôvodnom koryte došlo k zaklesnutiu hladiny podzemnej vody, redukcii plytkých zaplavených depresií a v nich nastupuje xeroséria sprevádzaná úbytkom výrazne vlhkomilných druhov (C. minimum, Oxyloma elegans, S. putris, Pseudotrichia rubiginosa).
Oblasť lužných lesov pri vyústení odpadového kanála do starého koryta Dunaja patrí k posledným zvyškom vnútrozemskej delty s výskytom vlhkého variantu mäkkého luhu as. Salici-Populetum myosotidetosum so zodpovedajúcou malakocenózou. Pôsobí tu spätné vzdutie vôd Dunaja, takže táto oblasť býva aj po prehradení Dunaja často zaplavovaná aj dva razy vo vegetačnom období. Kým pôvodná štruktúra bylinnej vegetácie je do značnej miery degradovaná (najmä prítomnosťou neofytu Aster lanceolata agg.), malakocenózy vykazujú štruktúru typickú pre takýto typ lužného lesa. V spoločenstve jednoznačne prevládajú autochtónne polyhygrofilné druhy Z. nitidus, C. minimum, P. rubiginosa, S. putris a O. elegans, ktoré tvoria často 90-95 % taxocenóz.
V úseku nedotknutom vodným dielom (Sap – Čičov) zvyšky lužných lesov v medzihrádzovom priestore bývajú zaplavované, prípadne dobre zásobované priesakovou vodou. Malakocenózy sa štruktúrou najviac podobajú pôvodným spoločenstvám najvlhších typov mäkkých lužných lesov, súdiac podľa prevahy polyhygrofilných druhov mäkkýšov, ktoré sú dobre adaptované na podmienky cyklického klimaxu. K typickým dominantám lesných malakocenóz patria najmä polyhygrofilné ulitníky C. minimum, Z. nitidus, S. putris, O. elegans a lesný hygrikolný druh V. crystallina (typickou ukážkou je napr. monitorovacia plocha Čičov-Starý les).
Vývoj výskytu rovnakonožcov (Oniscidae) dokumentuje najmä pomer dominancie eurytopného druhu Trachelipus rathkei tolerantného voči nižšej pôdnej vlhkosti, mezofilného druhu Hyloniscus riparius a hygrofilného druhu Porcellio scaber, ktorý sa na vlhkostnom gradiente výrazne mení. Na Dunajských krivinách boli zo 7 druhov zistených počas monitoringu (1993-1997) eudominantnými T. rathkei a H. riparius, mozaikovito rozšírený na zreteľne vlhkých miestach. Celkové druhové zloženie taxocenózy sa v rokoch po prehradení Dunaja podstatne nemenilo.
Na Bodíckej bráne bola zistená prítomnosť 7 druhov, v jednotlivých rokoch ich počet kolísal. Počas všetkých rokov bol eudominantný T. rathkei, v poslednom roku sledovania (1997) bol pozorovaný prudký pokles dominancie hygrofila P. scaber naznačujúci posun monitorovacej plochy k suchším stanovištiam. Výrub lesa nepochybne urýchlil tento proces v bezprostrednej blízkosti miesta, z ktorého sa v pravidelných intervaloch odoberali vzorky pôdnych článkonožcov.
V prvom roku monitoringu boli na Kráľovskej lúke zistené len H. riparius a eudominantný T. rathkei, v ďalších rokoch sa počet druhov zvýšil. Hodnota dominancie stenotopného hygrofila Porcellium collicola stúpala až na 46% (1997) a druhu T. rathkei prudko klesla z 98% (1993) na 13% v roku 1997; uvedené zmeny naznačujú zastavenie procesu vysychania. V Istragove počtom druhov chudobnejšie spoločenstvo s eudominantným druhom T. rathkei naznačovalo v rokoch 1993 až 1996 pomalý proces vysychania. Až v poslednom roku (1997) monitoringu tejto skupiny sa objavil hygrofilný Porcellium collicola s vysokým percentom dominancie, čo by mohlo naznačovať posun k zvýšenej vlhkosti ako možný pozitívny dôsledok umelých záplav.
V taxocenóze stonožiek - Chilopoda sa zmeny ktoré nastali po prehradení Dunaja odrazili na druhovom a kvantitatívnom zložení. Došlo k zmene počtu zistených druhov; okrem xenocénov vymizli Clinopodes flavidus, L. microps, ako nové boli zaznamenané Lithobius lapidicola a L. calcaratus patriace však do skupiny druhov menej náročných na pôdnu vlhkosť. Vzrástla dominancia eurytopného druhu Lithobius forficatus, pravdepodobne v dôsledku imigrácie po ústupe podmienok charakteristických pre lužný les, poklesla dominacia mezohygrofilného druhu Lithobius curtipes, charakteristického pre podunajské lužné lesy.
Na Dunajských krivinách v prvom roku po prehradení Dunaja (1993) zreteľne klesol počet zistených druhov na najnižšiu hodnotu (Tab. 5.1) a prudko vzrástla hodnota dominancie eurytopného druhu L. forficatus (z 21 na 49%), tolerantného voči silnému kolísaniu vlhkosti pôdy. Vysoká hodnota dominacie tohto druhu sa udržiavala aj v ďalších rokoch. Na druhej strane pozorujeme pokles dominacie aj u mezohygrofilného druhu L. curtipes, charakteristického pre podunajské lužné lesy (z 18 na 14% v roku 1993, v nasledujúcich rokoch až na 3-4%). Po prehradení Dunaja sa už nezistili Clinopodes flavidus, L. microps, L. lucifugus a L. cyrtopus (posledné dva sú xenocénne). Naopak, v rokoch po prehradení tu boli zaznamenané Lithobius lapidicola, L. pusillus a L. calcaratus patriace do skupiny druhov menej náročných na pôdnu vlhkosť. V roku 1995 sa počet zistených druhov zvýšil na 11 a hodnota dominancie eurytopného L. forficatus klesla na 27%. Je možné uvažovať o vzťahu uvedených zmien a umelou letnou záplavou. Kolísanie stupňa podobnosti (50-60%) spoločenstva stonožiek medzi rokmi 1993 až 1997 poukazuje na zatiaľ neustálené existenčné podmienky.
V Bodíckej bráne počet zistených druhov stonožiek po prehradení Dunaja klesol na 7 (Tab.5.2) a až v ďalších rokoch sa vyrovnal so stavom pred prehradením. Eurytopný L. forficatus bol po celé sledované obdobie dominantný. Hodnoty dominacie charakteristických druhov L. crassipes a L. curtipes kolísali a L. crassipes v niektorých rokoch nebol vôbec zistený. Dominacia hypogeického druhu Pachymerium ferrugineum sa po prehradení zvýšila na 20-30%. Uvedené zmeny v taxocenóze stonožiek poukazujú na posun tejto monitorovacej plochy k stanovištiam so suchším povrchom, čo potvrdzujú aj merania pôdnej vlhkosti.
Tab. 5.1 Počty jedincov zistených druhov stonožiek na lokalite Dunajské kriviny (MP č. 6)
Number of Chilopoda individuals and species on Dunajské kriviny monitoring plot (MP - 6)
Dobrohošť - Dunajské kriviny |
1991 |
1992 |
1993 |
1994 |
1995 |
1996 |
1997 |
|
Lithobius calcaratus |
- |
- |
- |
1 |
- |
- |
- |
|
L. cyrtopus |
- |
1 |
- |
- |
- |
- |
- |
|
L. erythrocephalus |
9 |
11 |
1 |
3 |
10 |
5 |
4 |
|
L. forficatus |
23 |
21 |
49 |
64 |
27 |
43 |
29 |
|
L. lapidicola |
- |
- |
- |
- |
2 |
2 |
- |
|
L. luteus |
- |
- |
- |
- |
- |
- |
2 |
|
L. lucifugus |
- |
1 |
- |
- |
- |
- |
- |
|
L. mutabilis |
5 |
3 |
7 |
8 |
5 |
- |
10 |
|
L. muticus |
- |
- |
2 |
- |
- |
- |
1 |
|
L. aeruginosus |
9 |
13 |
12 |
9 |
15 |
6 |
3 |
|
L. austriacus |
- |
- |
- |
1 |
1 |
- |
8 |
|
L. crassipes |
7 |
4 |
7 |
4 |
4 |
4 |
4 |
|
L. curtipes |
27 |
18 |
14 |
4 |
4 |
7 |
3 |
|
L. microps |
3 |
- |
- |
- |
- |
- |
- |
|
Clinopodes flavidus |
- |
1 |
- |
- |
- |
- |
- |
|
Pachymerium ferrugineum |
8 |
4 |
4 |
5 |
26 |
12 |
9 |
|
P. tristanicum |
- |
2 |
- |
1 |
4 |
1 |
4 |
|
Strigamia acuminata |
- |
- |
- |
- |
- |
- |
1 |
|
Geophilus flavus |
9 |
4 |
- |
- |
1 |
4 |
19 |
|
Počet druhov – Number of species |
9 |
12 |
8 |
10 |
11 |
9 |
13 |
Tab. 5.2 Počty jedincov zistených druhov stonožiek na lokalite Bodícka brána (MP č. 9)
Number of Chilopoda individuals and species on Bodícka brána monitoring plot (MP - 9)
|
Bodíky - Bodícka brána |
1991 |
1992 |
1993 |
1994 |
1995 |
1996 |
1997 |
|
Lithobius agilis |
- |
8 |
- |
- |
- |
- |
- |
|
L. erythrocephalus |
11 |
13 |
4 |
4 |
- |
3 |
20 |
|
L. forficatus |
43 |
34 |
56 |
52 |
19 |
21 |
32 |
|
L. lapidicola |
- |
- |
- |
- |
- |
- |
1 |
|
L. mutabilis |
1 |
5 |
16 |
12 |
5 |
12 |
5 |
|
L. muticus |
- |
2 |
- |
- |
- |
- |
- |
|
L. piceus |
1 |
- |
- |
- |
- |
- |
- |
|
L. aeruginosus |
- |
8 |
12 |
20 |
2 |
1 |
- |
|
L. austriacus |
- |
- |
- |
- |
- |
2 |
- |
|
L. crassipes |
17 |
- |
2 |
- |
19 |
5 |
- |
|
L. curtipes |
18 |
18 |
2 |
4 |
10 |
24 |
5 |
|
L. microps |
1 |
- |
- |
- |
- |
- |
- |
|
Pachymerium ferrugineum |
6 |
5 |
- |
4 |
34 |
21 |
20 |
|
P. tristanicum |
- |
- |
- |
- |
3 |
1 |
1 |
|
Strigamia acuminata |
1 |
5 |
- |
- |
- |
- |
- |
|
Geophilus flavus |
1 |
- |
6 |
4 |
- |
- |
11 |
|
Schendyla nemorensis |
- |
- |
- |
- |
4 |
10 |
4 |
|
Počet druhov – Number of species |
10 |
9 |
7 |
7 |
8 |
10 |
9 |
Spoločenstvo stonožiek Kráľovskej lúky pred prehradením Dunaja zaraďujeme medzi najbohatšie. Za obdobie rokov 1991 – 1997 tu bolo zistených 17 druhov, po prehradení Dunaja počet druhov zreteľne klesol. Taxocenózu stonožiek tvorilo 13 druhov (Tab. 5.3), medzi ktorými bol pre tento typ stanovišťa charakteristický ripikolný a hygrofilný Lamyctes emarginatus a hygrofilné druhy L. agilis a L. microps. Eudominantným druhom bol L. emarginatus, dominantnými L. aeruginosus a L. curtipes. Po roku 1993 tu už nebol zistený náročnejší hygrofilný druh L. agilis a lesný druh charakteristický pre vyššie polohy L. cyrtopus, ktorý je však pre lužné lesy cudzím prvkom a na toto územie sa pravdepodobne dostával počas povodní splavením. V spoločenstve stonožiek po r. 1993 tiež chýba L. microps. Hypogeické druhy Geophilus flavus, P. ferrugineum a Strigamia acuminata, zistené pred prehradením Dunaja, sa po dočasnej absencii opätovne vo vzorkách presevu objavili až v r. 1997 (asi ako výsledok simulovaných záplavam). Zmeny spoločenstva stonožiek sa neukončili ani v r. 1997, kedy tu boli po prvýkrát zistené Lithobius pelidnus a Pachymerium tristanicum. Zmeny v druhovej skladbe spoločenstva signalizujú neustálenosť podmienok tejto lokality. Dominantné až eudominantné zastúpenie na lokalite si udržiavali eurytopný L. forficatus a mezohygrofilné druhy L. aeruginosus, L. crassipes a L. curtipes. Ripikolný druh L. emarginatus zaznamenal pokles dominancie v rokoch 1993-1995, vzostup v r. 1996 a absenciu v r. 1997. Kolísanie hodnoty dominancie tohoto druhu bolo odrazom vtedajšieho vysychania pobrežného pásma mŕtveho ramena, ležiaceho bližšie k hrádzi. Nestabilné pomery na monitorovacej ploche sa v plnom rozsahu preniesli na taxocenózu stonožiek, ktorej podobnosť medzi rokmi sledovania mala najnižšiu hodnotu pri porovnaní rokov 1992-1993.
Tab. 5.3 Počty jedincov zistených druhov stonožiek na lokalite Kráľovská lúka (MP č. 10)
Number of Chilopoda individuals and species on Kráľovská lúka monitoring plot (MP - 10)
Bodíky - Kráľovská lúka |
1991 |
1992 |
1993 |
1994 |
1995 |
1996 |
1997 |
|
Lamyctes emarginatus |
21 |
13 |
2 |
4 |
5 |
15 |
- |
|
Lithobius agilis |
19 |
- |
- |
- |
- |
- |
- |
|
L. cyrtopus |
- |
1 |
- |
- |
- |
- |
- |
|
L. erythrocephalus |
3 |
8 |
4 |
11 |
7 |
5 |
11 |
|
L. forficatus |
5 |
11 |
9 |
24 |
20 |
29 |
12 |
|
L. mutabilis |
16 |
18 |
41 |
31 |
17 |
17 |
25 |
|
L. muticus |
3 |
2 |
- |
3 |
- |
- |
6 |
|
L. pelidnus |
- |
- |
- |
- |
- |
- |
1 |
|
L. tricuspis |
8 |
2 |
- |
- |
- |
- |
- |
|
L. aeruginosus |
3 |
20 |
15 |
14 |
4 |
9 |
- |
|
L. crassipes |
- |
- |
25 |
4 |
19 |
6 |
4 |
|
L. curtipes |
- |
11 |
4 |
7 |
24 |
15 |
31 |
|
L. microps |
8 |
5 |
- |
- |
- |
- |
- |
|
Pachymerium ferrugineum |
- |
3 |
- |
- |
- |
- |
1 |
|
P. tristanicum |
- |
- |
- |
- |
- |
- |
3 |
|
Strigamia acuminata |
13 |
- |
- |
- |
4 |
1 |
- |
|
Geophilus flavus |
- |
2 |
- |
- |
- |
- |
4 |
|
Počet druhov – Number of species |
10 |
12 |
7 |
8 |
8 |
8 |
10 |
Na Istragove bolo v taxocenóze stonožiek za sledované obdobie 1991-1997 zistených 15 druhov, z ktorých medzi eudominantné patrili eurytopné L. forficatus a L. mutabilis. Z typických mezohygrofilných druhov boli vysokým percentom zastúpené L. aeruginosus, L. crassipes, L. curtipes a hypogeicky žijúci druh P. ferrugineum. Na lokalite pred prehradením Dunaja nechýbal ani hygrofilný druh L. agilis, zistený v rokoch 1991 a 1992. Signálom vysychania monitorovacej plochy je počínajúc rokom 1993 absencia hygrofilného druhu L. agilis, rast hodnôt dominancie vlhkostne tolerantného druhu L. forficatus. Počty zistených druhov v rokoch po prehradení sú uvedené v Tab.5.4. Dominantné postavenie počas rokov sledovania si udržiavajú eurytopný L. forficatus a mezohygrofilné druhy L. curtipes a L. crassipes. Zvýšenie hodnoty dominancie hypogeického druhu P. ferrugineum z 10% až na 31% v roku 1996, preferujúceho piesčité, no nie mokré či zabahnené stanovištia, tiež naznačuje posun lokality k suchším stanovištiam. Najvýraznejšie zmeny podobnosti taxocenóz stonožiek sú medzi rokmi pred prehradením a posledným rokom (1997) monitorovania (33%, 41%). Sú tiež potvrdením pomalého posunu k suchšiemu typu stanovišťa. čoho dôkazom je zmena druhového zastúpenia stonožiek a prítomnosť druhov tolerantných voči nižšiemu stupňu vlhkosti.
Tab. 5.4 Počty jedincov zistených druhov stonožiek na lokalite Istragov (MP č. 14)
Number of Chilopoda individuals and species on Istragov monitoring plot (MP - 14)
Istragov |
1991 |
1992 |
1993 |
1994 |
1995 |
1996 |
1997 |
|
Lithobius agilis |
2 |
3 |
- |
- |
- |
- |
- |
|
L. erythrocephalus |
- |
7 |
2 |
12 |
8 |
4 |
6 |
|
L. forficatus |
10 |
21 |
23 |
34 |
15 |
24 |
28 |
|
L. latro |
- |
- |
- |
- |
1 |
- |
- |
|
L. mutabilis |
- |
32 |
8 |
9 |
14 |
7 |
8 |
|
L. pelidnus |
- |
- |
- |
- |
- |
- |
4 |
|
L. aeruginosus |
20 |
14 |
6 |
4 |
- |
- |
- |
|
L. biunguiculatus |
- |
3 |
- |
- |
- |
- |
- |
|
L. crassipes |
2 |
- |
36 |
7 |
14 |
22 |
10 |
|
L. curtipes |
60 |
7 |
8 |
21 |
5 |
6 |
2 |
|
C. linearis |
- |
- |
2 |
2 |
- |
- |
- |
|
Pachymerium ferrugineum |
6 |
10 |
12 |
10 |
27 |
31 |
26 |
|
P. tristanicum |
- |
- |
- |
- |
3 |
4 |
6 |
|
Geophilus flavus |
- |
- |
- |
- |
- |
- |
6 |
|
Schendyla nemorensis |
- |
|
- |
- |
- |
- |
2 |
|
Počet druhov – Number of species |
6 |
8 |
8 |
8 |
8 |
7 |
10 |
Zo bzdôch (Heteroptera) pre posúdenie stavu a zmien sú indikačne významné predovšetkým druhy žijúce v listovej opadanke a v povrchových vrstvách pôdy, ktoré citlivo reagujú na zmeny charakteru lužných lesov, najmä na vlhkostné pomery, zásahy do lesných spoločenstiev, ich deštrukciu, výrub a následnú aridizáciu a ruderalizáciu. Na monitorovacích plochách bolo zistených 85 druhov s celkovou priemernou abundanciou PA = 6.23 ex.m-2 (Štepanovičová, Degma 1999). Taxocenózy epigeických Heteroptera celého záujmového územia sa vyznačujú vysokým stupňom heterogenity ovplyvnenej nízkym počtom konštantných a eukonštantných druhov (10), všetky ostatné druhy predstavujú v taxocenózach akcesorické a akcidentálne prvky. Najväčší počet týchto druhov sa pochopiteľne vyskytoval v najsuchšom variante lužného lesa, kam najmä v čase hibernácie migrovali viaceré druhy z blízkeho xerotermofilného spoločenstva dunajskej hlošiny. Z aspektu kvalitatívnej štruktúry sa týmto dvom cenózam najviac približuje taxocenóza bzdôch na lokalite Dunajské kriviny (PA = 9.33 ex.m-2), kde sa vyskytli vyššie počty druhov najmä po r. 1995 ako dôsledok ruderalizácie. Kvantitatívne zastúpenie bzdôch na tejto lokalite dosiahlo najvyššiu mieru podobnosti s cenózou v Bodíckej bráne (10.63 ex.m–2); na oboch lokalitách boli hodnoty PA súčasne najvyššie zo všetkých v priebehu celého obdobia výskumu. Zo zistených druhov však iba 17 možno hodnotiť ako druhy pre epigeón lužného lesa charakteristické (cenobiontné alebo cenofilné). Pre indikáciu zmien sú použité iba tie hygrofilné resp. mezohygrofilné druhy, ktoré kvantitatívnymi ukazovateľmi (PA, dominancie, konštantnosti) vyhovujú požiadavkám na indikačne významné druhy: Drymus brunneus, Drymus ryeii, Scolopostethus affinis, Scolopostethus thomsoni a Legnotus limbosus. Pri posudzovaní zmien taxocenózy v spojení so zmenami, ku ktorým dochádzalo v priebehu výstavby vodného diela, sa vychádza z ekologických nárokov zistených druhov a ich vzťahu k podmienkam habitatu. Indikačné schopnosti charakteristických druhov sa výrazne prejavili najmä v úseku derivácie, kde dosiahlo ich kvantitatívne zastúpenie až 74.88%. Najvyššie hodnoty dominancie charakteristických druhov sa zaznamenali v r. 1996 a 1997, kedy sa pomery v epigeóne lužných lesov konsolidovali a taxocenózy bzdôch dosiahli stav podobný ako v roku 1991 (Tab. 5.5).
Výrazný pokles v hodnotách dominancie charakteristických druhov bol zaznamenaný v r. 1993 – 1995 na lokalite v Dobrohošti. V tomto období sa prejavil negatívny vplyv výrubu časti lužného lesa a jeho postupné vysychanie. V dôsledku postupujúcej aridizácie a ruderalizácie lužného lesa začali do lesných spoločenstiev prenikať z okolitých biotopov na vlhkosť menej náročné euryekné druhy bzdôch, čo sa popri poklese dominancie charakteristických druhov prejavilo aj na ich vyššom kvalitatívnom zastúpení (r. 1995 – 15 spp.).
Výrazné rozdiely v kvalitatívnej i kvantitatívnej štruktúre taxocenóz Heteroptera medzi monitorovacími plochami Bodícka brána, Kráľovská lúka a Istragov sa prejavili predovšetkým nízkymi hodnotami priemernej abundancie (Kráľovská lúka za celé obdobie 1.97 ex.m-2, Istragov 3.09 ex.m–2) niekoľkonásobne nižšími ako na dvoch predchádzajúcich plochách. Na základe 7 ročného výskumu sa ukázalo že na Kráľovskej lúke v pôdnej depresii vlhkosť listovej opadanky a pôdy prekračovala hranice tolerancie aj hygrofilných suchozemských bzdôch, ktoré sa tu preto vyskytovali len v malom počte jedincov.
Tab. 5.5 Hodnoty priemernej abundancie (A.m-2 ), celkový počet druhov (spp.) a dominancia charakteristických druhov (D) bzdôch v oblasti inundácie v r. 1991 – 1997
Average abundace (A.m-2), number of species (spp.) and dominance of characteristic species (D) of bugs in the floodplain in 1991 - 1997
|
Rok – Year / MP |
MP 6 |
MP 9 |
MP 10 |
MP 14 |
|
A.m-2 1991 spp. D |
8.18 4 69.02 |
17.18 16 83.92 |
1.33 3 50.00 |
3.50 9 85.72 |
|
A.m-2 1992 spp. D |
8.42 7 88.14 |
8.83 4 98.22 |
0.50 3 37.50 |
2.50 5 80.00 |
|
A.m-2 1993 spp. D |
5.33 9 62.62 |
7.00 11 82.14 |
1.50 3 50.00 |
5.16 6 85.72 |
|
A.m-2 1994 spp. D |
7.33 5 68.00 |
6.66 5 92.50 |
2.16 6 84.62 |
2.50 5 86.66 |
|
A.m-2 1995 spp. D |
9.66 15 54.40 |
10.53 9 80.63 |
1.83 8 36.36 |
2.50 7 66.66 |
|
A.m-2 1996 spp. D |
9.53 11 80.60 |
7.33 8 84.09 |
4.60 4 73.91 |
4.50 9 64.67 |
|
A.m-2 1997 spp. D |
8.14 8 77.17 |
4.42 10 87.10 |
1.87 7 100.00 |
1.00 3 84.80 |
|
Vysvetlivky: |
N – výskum sa nekonal |
MP – Monitorovacia plocha |
| MP 6 – Dobrohošť – Dunajské kriviny, | MP 9 – Bodícka brána, | |
| MP 10 – Kráľovská lúka, | MP 14 – Istragov, | |
| MP 18 – Kľúčovec – Sporná sihoť, | MP 23 – Čičov – Starý les |
Na posúdenie rozdielov vlhkostných pomerov listovej opadanky lužných lesov sa ukázalo vhodným využitie údajov o výskyte charakteristických dominantných druhov D. brunneus a L. limbosus s dobrými indikačnými vlastnosťami (Štepanovičová 1997) a rozdielmi v ich kvantitatívnom zastúpení (Tab. 5.6). Odlišná ekologická charakteristika týchto dvoch druhov (D. brunneus je hygrofil, typický obyvateľ listovej opadanky lužných lesov, L. limbosus mezofil) umožňuje na základe ich výskytu a populačnej hustoty charakterizovať trvalé aj prechodné zmeny vlhkostných pomerov ich habitatov. Maximálne hodnoty abundancie v Dobrohošti dosahovali 10.50 ex.m-2 v r. 1992 a na lokalite Bodícka brána 7.54 ex.m-2 v r. 1991 t. j. pred nástupom rozsiahlejších stavebných prác a vodohospodárskych úprav. Náhly pokles, ktorý sa v ich dôsledku prejavil v r. 1993 sa v neskorších rokoch, v súvislosti so simulovanými záplavami, postupne vyrovnal. Populácie L. limbosus sa vyskytovali pravidelne iba v najsuchšom variante lužného lesa mimo úseku inundačného územia (Kopáč, Ostrovné lúčky) a maximálne hodnoty abundancie dosiahli na oboch týchto plochách v r. 1995. Na ostatných monitorovacích plochách bol výskyt druhu sporadický, nepravidelný a jeho nízka až podpriemerná abundancia resp. absencia nemajú takú výpovednú hodnotu, ktorá by svedčila o zmenách vlhkostných pomerov lužných lesov, najmä o poklese vlhkosti a zreteľnom vysychaní.
Tab. 5.6 Hodnoty priemernej abundancie (ex.m-2) druhov Drymus brunneus (A) a Legnotus limbosus (B) na monitorovacích plochách (MP)
Average abundance (ex.m-2 ) of species Drymus brunneus (A) and Legnotus limbosus (B) on monitoring plots (MP)
|
Rok – Year / MP |
MP 6 |
MP 9 |
MP 10 |
MP 14 |
MP 18 |
MP 23 |
|
1991 A B |
4.81 - |
7.54 - |
0.13 - |
0.13 - |
0.13 - |
N
|
|
1992 A B |
10.50 0.14 |
3.50 - |
- - |
1.12 - |
5.32 - |
N
|
|
1993 A B |
2.38 0.56 |
0.28 - |
0.42 - |
2.24 - |
2.70 0.42 |
3.64 0. 12 |
|
1994 A B |
5.12 - |
3.52 - |
0.46 - |
0.80 - |
1.12 1.44 |
0.96 1.76 |
|
1995 A B |
3.16 0.28 |
6.83 0.50 |
0.16 - |
0.83 0.66 |
0.83 0.33 |
0.80 0.49 |
|
1996 A B |
2.73 2.83 |
1.33 - |
1.05 1.16 |
0.83 0.33 |
0.16 - |
0.69 0.16 |
|
1997 A B |
5.33 0.73 |
1.75 0.52 |
0.40 - |
0.16 - |
0.85 - |
3.50 - |
Vysvetlivky pozri Tab. 5.5
Rozdiely v hodnotách priemernej abundancie druhov D. brunneus a L. limbosus sú výrazné aj pri ich sumarizácii do troch skupín lesných spoločenstiev. V najsuchšom variante lužného lesa bola priemerná abundancia D. brunneus iba 0.13 ex.m-2, L. limbosus 2.60 ex.m-2. Na lokalitách úseku derivácie dosiahla hodnota priemernej abundancie D. brunneus 2.50 ex.m-2, L. limbosus iba 0.31 ex.m-2. Na lokalitách pod miestom návratu vôd Dunaja do pôvodného koryta bola abundancia D. brunneus 1.72 ex.m-2 a L. limbosus 0.41 ex.m-2. Rozdiely v hustote populácií uvedených druhov vhodne indikujú lužné lesy a habitat ich listovej opadanky s najnižším, najvyšším a priemerným stupňom vlhkosti.
Na exaktný dôkaz vplyvu niektorých faktorov na formovanie taxocenóz bzdôch v epigeóne lužných lesov sa použila metóda kanonickej korešpondenčnej analýzy, ktorá ukázala (Štepanovičová, Degma 1999), že z 9 testovaných faktorov majú pre formovanie taxocenóz epigeických bzdôch význam iba 4 gradientové premenné – vlhkosť pôdy a listovej opadanky, pH pôdy, obsah CaCO3 v pôde a priemerná teplota ovzdušia. Vedúcim faktorom ovplyvňujúcim výskyt bzdôch v epigeóne lužného lesa je vhodný stupeň vlhkosti. Najužšiu afinitu k tomuto faktoru majú charakteristické druhy D. brunneus, S. affinis, S. thomsoni a tri ďalšie hygrofilné druhy, S. pilosus, H. saturejae a E. testudinarius.
Ukázalo sa, že dostatočný stupeň vlhkosti listovej opadanky, ktorá je dominantným faktorom pre existenciu populácií charakteristických druhov bzdôch, pretrvával aj v posledných rokoch monitoringu tejto skupiny (1996 a 1997). Z výsledkov 7 ročného výskumu súčasne vyplýva, že na rozdiel od pozitívne pôsobiacej dostatočnej vlhkosti listovej opadanky lužných lesov, ovplyvňovanej hladinou podzemnej vody, povrchová voda, ktorej stav je najvyšší v čase záplav, spôsobuje kratšie i dlhšie trvajúci pokles kvantitatívneho ale aj kvalitatívneho zastúpenia bzdôch, čo sa prejavuje nižšími hodnotami abundancie charakteristických druhov a poklesom druhovej diverzity taxocenóz epigeických bzdôch.
Na definovanie optimálneho stavu podmienok pre existenciu populácií autochtonných epigeických bzdôch v listovej opadanke a v povrchových vrstvách pôdy lužných lesov Podunajska možno využiť uvedené údaje o kvalitatívnej aj kvantitatívnej štruktúre ich taxocenóz, z ktorých jednoznačne vyplýva požiadavka na vhodné vlhkostné pomery, ovplyvňované najmä výškou hladiny podzemnej vody. Dôkazom toho, že optimálny stav pre bzdochy v lužných lesoch pretrváva, je predovšetkým vysoká hustota populácií charakteristických hygrofilných druhov, ktorých dominancia dosiahla na celom záujmovom území 68% a v úseku derivácie, kde sú vlhkostné pomery priaznivejšie, až 74.88%.
V prirodzených spoločenstvách bystruškovitých (Coleoptera: Carabidae) v lužných lesoch sa počet druhov pohybuje väčšinou od 22 do 35, zriedkavo dosahuje až 50 druhov. Počet druhov klesajúci pod 15 je už príznakom značne degradovaných spoločenstiev (ak výsledok nie je ovplyvnený metodickou chybou).
Pri celkovom hodnotení pre prirodzené spoločenstvá bystruškovitých v ekosystémoch v najvlhších častiach vnútrozemskej delty Dunaja v podmienkach cyklického katastrofického klimaxu sú charakteristické hodnoty kumulatívnej abundancie v medziach cca 80-130 ex/pasca/rok. Pokles pod túto hodnotu, pokiaľ nie je spôsobený dlhodobým zaplavením, možno interpretovať ako známku degradácie. Medzné najnižšie hodnoty (stanovené aj na základe materiálov z iných lokalít) v silne degradovaných spoločenstvách sa pohybujú v rozmedzí 5-20 ex/pasca/rok. Vyššie hodnoty ako 130 ex/pasca/rok majú rôznu interpretáciu. Pokiaľ sa vyskytujú jednorázovo a sú spôsobené prudkým zvýšením početnosti druhov Pterostichus melanarius a/alebo Pterostichus niger možno ich chápať ako známku prechodného zníženia vlhkosti na príslušnej lokalite. Prudké zvýšenie zastúpenia P. niger sprevádza počiatočne fázy sukcesie na dlhodobo zaplavených plochách. Pokiaľ zmena kumulatívnej abundancie bola spôsobená zvýšením početnosti niektorých hygrofilných druhov (Agonum moestum, Pterostichus anthracinus, P. nigrita, Bembidion mannerheimi, B. biguttatum), je ju treba chápať ako známku straty charakteru spoločenstva žijúceho v podmienkach cyklického klimaxu typického pre podunajské lužné lesy a ich priblíženie sa podmienkam v prirodzených lužných lesoch zaplavovaných stagnujúcou vodou. V takýchto prípadoch sú najčastejšie hodnoty kumulatívne abundancie okolo 200-300 ex/pasca/rok, výnimočne až 400-500 ex/pasca/rok.
Zmena zastúpenia ekologických skupín druhov je z hľadiska potrieb bioindikácie zmien spoločenstiev v lužných lesoch najvýznamnejším kritériom. Zvlášť významný je pomer zastúpenia druhov s rozličnými vlhkostnými nárokmi. U bystruškovitých presné experimentálne zistené hodnoty hygropreferenda pre jednotlivé druhy nie sú známe. Na základe komparácie a terénnych pozorovaní však možno druhy semikvantitatívne klasifikovať do ôsmich skupín. Ich zastúpením je možné jednoduchým spôsobom kvantifikovať prebiehajúce zmeny a použiť ho na metódu priamej ordinácie spoločenstiev.
Ďalším významným kritériom je pomer druhov vyžadujúcich trvalé zatienenie drevinnou vegetáciou, druhov indiferentných k zatieneniu a druhov vyžadujúcich nelesné ekosystémy.
Idealizovanú štruktúru spoločenstva, zodpovedajúceho týmto podmienkam zachytáva Tab. 2.1 a Tab. 2.2. Tieto spoločenstvá sa vyznačujú vysokým podielom silne hygrofilných druhov (v tabuľkách stupne 6-8) a absenciou alebo nízkym zastúpením hygrofilných druhov Pterostichus. anthracinus, P. nigrita, Bembidion mannerheimi a B. biguttatum. Pozíciu týchto spoločenstiev v systéme viacerých referenčných spoločenstiev ukazujú priame ordinácie urobené na základe nárokov bystruškovitých na vlhkosť a vegetačný kryt.
Výskyt hygrofilných druhov bystruškovitých v Dunajských krivinách v roku 1993 výrazne klesol. Niektoré druhy sa po roku 1993 prestali vyskytovať, naopak zvýšil sa počet na vlhkosť menej náročných druhov, najmä Carabus granulatus (1993: 21,59%) a Pterostichus niger (1992: 25,52%). Ich zvýšený výskyt trval do roku 1995 kedy opäť poklesol a v roku 1997 tieto druhy neboli zistené. V rokoch 1994 sa začala invázia xenocénneho druhu Pseudophonus rufipes, ktorá vyvrcholila v roku 1995 (17,12% jedincov). Priebeh jeho invázie časovo koincidoval s ojedinelým výskytom viacerých xerofilných druhov. Ich najvýraznejším predstaviteľmi boli Licinus depressus a viaceré druhy rodu Amara. Počas celého obdobia bol relatívne najstabilnejší výskyt na vlhkosť menej náročného druhu Stomis pumicatus (okolo 10%). Priebeh zmien taxocenózy bystruškovitých od východiskového stavu v roku 1989 k stavu v roku 1997 charakterizuje kontinuálny pokles hodnôt indexov podobnosti celoročných zberov z rokov 1990-1997 so zberom v roku 1989 (z 43-62% na 16-30%). Z úseku derivácie bolo spoločenstvo bystruškovitých najviac postihnuté zmenami v hornej časti (monitorovacia plocha Dunajské kriviny, MP-6).
V roku 1993 sa na lokalite Bodícka brána prudko zvýšilo absolútne i relatívne zastúpenie tolerantnejších druhov C. granulatus (11,11%), Pterostichus melanarius (25,90%), P. niger (24,26%) a Stomis pumicatus (3,89%). Súčasne kleslo zastúpenie všetkých vlhkostne náročnejších druhov. V rokoch 1994-1996 abundancia prevažnej väčšiny druhov výrazne poklesla. Odrazilo sa to znížením kumulatívnej abundancie približne na 1/3-1/2 stavu z roku 1993. Najviac boli postihnuté najnáročnejšie druhy, najmä P. assimilis, Clivina fossor, B. dentelum a B. sodalis. Postihnuté neboli len niektoré menej náročné alebo tolerantné druhy, najmä P. strenuus a A. flavipes a O. obscurus pri ktorých sa v jednotlivých rokoch naopak abundancia mierne zvýšila. Pri týchto druhoch hrá významnú úlohu aj okolnosť, že ide o somaticky malé druhy. Zmeny abundancie jednotlivých druhov preto zrejme odrážajú nielen samotné zmeny hydrologického režimu, ale sprostredkovane aj zmeny potravnej ponuky. Najhorší stav bol v roku 1995. Od roku 1993 nastáva na lokalite invázia euryekného xenocénneho druhu Trechus quadristriatus, ktorú možno považovať za výraznú degradatívnu zmenu. Pomer zastúpenia jednotlivých druhov naznačuje, že v roku 1997 nastalo mierne zlepšenie vlhkostných pomerov na tejto lokalite. Trend zmien je mierny.
Na Kráľovskej lúke čiastočne už v roku 1992, ale vo výraznej podobe až v roku 1993, sa zvýšilo zastúpenie tolerantnejších alebo vlhkostne menej náročných druhov C. granulatus (35,6 a 22,5%), P. melanarius (6,5 a 14,2%), P. niger (13,5 a 22,3%). Vo väčšom počte sa objavil ako nový druh na lokalite vlhkostne menej náročný Epaphius secalis. Po poklese v r. 1991 sa opäť absolútne zvýšilo aj zastúpenie dvoch menej tolerantných vlhkomilných druhov Platynus assimile a Patrobus atrorufus, ktorého zastúpenie však relatívne pokleslo. Pri obnovení zberov, prerušených v rokoch 1994-5, bol zistený výrazný pokles počtu jedincov (z 1648 v r. 1993 na 636 v r. 1996 a 396 v r. 1997). V spoločenstve v r. 1996 a 1997 dominovali len tri tolerantné druhy P. strenuus (30,1%, resp. 36,36%), C. granulatus (16,82%, resp.19,19%) a O. obscurus (v r. 1993 10,1%). Relatívne i absolútne zastúpenie týchto troch druhov výrazne ovplyvnilo aj zvýšenie indexov podobnosti celoročných zberov z r. 1996 a 1997 v porovnaní so zberom v r. 1987, a to napriek nižšiemu zastúpeniu viacerých vlhkomilných druhov. V roku 1996 sa prejavila aj invázia xenocénneho T. quadristriatus, v roku 1997 však tento druh čiastočne ustúpil. Index druhovej podobnosti k roku 1987 vykazoval stabilizovaný trend s malými fluktuáciami. Indexy proporcionálnej podobnosti a podobnosti abundancie v závislosti na opísaných zmenách zastúpenia druhov veľmi prudko poklesli v rokoch 1989, 1992 a 1993. V rámci lokality sa v závislosti na zmenách vlhkosti a na drobných terénnych výškových rozdieloch v nadmorskej výške (rádovo 50-60 cm) výrazne menilo priestorové rozšírenie druhov (Šustek 1995). Po vysušení lokality a ústupe silne vlhkomilných druhov, tolerantnejšie druhy zaujali ich miesto v nižšie položených častiach, pričom vyvýšené miesta boli výrazne menej osídlené.
Na Istragove, najnižšie ležiacej monitorovacej ploche úseku derivácie, v roku 1989 výrazne prevažovali dva drobné vlhkomilné druhy bystruškovitých A. flavipes (47,7%) a B. femoratum (15,36%), výrazné zastúpenie mali aj ďalšie vlhkomilné druhy P. assimile (15,5%), P. strenuus (5,7%), C. granulatus (3,6%), P. atrorufus (2,4%). Ekologicky významná bola prítomnosť druhov Europhilus fuliginosus a E. micans. Vysoké zastúpenie vlhkostne menej náročných druhov ešte pred podstatnými zmenami hydrologického režimu odráža prevahu silne piesčitej pôdy na veľkej časti sledovanej plochy. V rokoch 1990-1992 klesla kumulatívna abundancia na polovicu. Najväčší podiel na tom mal pokles zastúpenia A. flavipes a B. femoratum. Približne na 2/3 až 1/2 klesla aj abundancia takmer všetkých ďalších druhov. Už v r. 1991 nastalo prudké zvýšenie zastúpenia P. niger, ktoré sa na ďalších lokalitách dostavilo až o dva roky neskôr. Počas rokov 1990 - 1992 sa vytvoril dočasne stabilizovaný stav, ktorý sa zmenil až v r. 1993 silným zvýšením abundancie C. granulatus (z 58 jedincov v r. 1992 na 320 jedincov v r. 1993), P. atrorufus (z 11 na 211 jedincov) a P. strenuus (z 24 na 222 jedincov). Súčasne sa zvýšila aj kumulatívna abundancia viac ako dvojnásobne a v r. 1995 klesla približne na úroveň r. 1992. Súčasne absolútne i relatívne opäť prudko vzrástlo zastúpenie Asaphidion flavipes, kým zastúpenie väčšiny ostatných druhov kleslo. Absolútne sa výrazne zvýšilo iba zastúpenie vlhkostne málo náročného E. secalis. V rokoch 1996-1997 sa pomer zastúpenia druhov priblížil stavu v r. 1990-1991, ďalej však klesala kumulatívna abundancia. Záplava v r. 1997 podporená spätným vzdutím vody v bývalom hlavnom koryte umožnila dokonca výskyt niektorých paludikolných druhov (Agonum moestum, Badister sodalis, E micans).
Fytofágne nosáčiky (Curculionoidae) predstavujú ekologické dominanty v rámci spoločenstiev chrobákov v lesných, lúčnych i mokraďových ekosystémoch. Sú primárnymi konzumentami v rade konvergentov a sú úzko naviazané na príslušné rastlinné spoločenstvo, citlivo reagujú na zmeny fytocenózy. Ich štúdium vo vegetačnom leme dunajských ramien prebiehalo od obdobia prehradenia Dunaja do r. 1997 na 6 monitorovacích plochách, kde bolo zistených 113 druhov. Ich prehľad s uvedením základných ekologických charakteristík je v Tab. 5.7.
V minulosti na monitorovacej ploche Dobrohošť - Dunajské kriviny rástli fytocenózy, ktoré patrili do asociácií Phalaridetum arundinaceae, Phragmitetum communis a Rorippo - Agrostietum stoloniferae. Rameno nachádzajúce sa na okraji lužného lesa je však od roku 1993 bez vody, postupne došlo k ruderalizácii jeho dna. Meniacu sa taxocenózu nosáčikov tvorilo 51 druhov zistených počas rokov 1993-1997, no iba Phyllobius sinuatus, Polydrusus sericeus, Gymnetron tetrum boli zastúpené vo vzorkách zo všetkých rokov. Hodnoty priemernej abundancie nosáčikov signifikantne klesali k roku 1997. Výrazný ústup vlhkomilných druhov z lokality pokračoval aj v poslednom roku monitoringu (1997). Z druhov náročnejších na vlhkosť sa v r. 1997 ešte vyskytovali Chlorophanus viridis, Phyllobius pomaceus a Lepyrus armatus. Ide o druhy zo susedného lužného lesa, ktoré dokážu tolerovať vysychanie biotopu. Na monitorovacej ploche sa prestali vyskytovať druhy Poophagus sisymbrii, Chlorophanus graminicola, Hypera rumicis. Zvýšil sa percentuálny podiel xerofilných druhov a druhov charakteristických pre ruderálnu vegetáciu (napr. Apion brevirostre, Cionus olivieri, Gymnetron asellus, G. bipustulatum) . Lokalitu možno už od r. 1994 hodnotiť ako silne narušenú. Zreteľný bol totálny ústup hygrofilnejších druhov rastlín a ich fytofágov.
Slepé a trvalo zatienené rameno na lokalite Bodíky - Bodícka brána je charakteristické vysokým stavom vody počas celého roku. Z rastlinných spoločenstiev prevládajú asociácie Rorippo amphibiae - Oenanthetum aquaticae, Rorippa amphibia - Agrostis stolonifera as., Myriophyllum spicatum as. a Lemno - Spirodeletum (frag.), ktoré prevláda na vodnej hladine ramena. Počas monitoringu bola zistená prezencia 32 druhov nosáčikov, z toho ale iba dva charakteristické druhy (cenobionty) P. sisymbrii a Tanysphyrus lemnae sa vyskytovali počas všetkých rokov. Spoločenstvo je kvantitatívne bohaté a zaznamenali sme tu najvyššie hodnoty priemernej abundancie počas celého sledovaného obdobia. Lokalitu možno hodnotiť ako veľmi zachovalú. Vzhľadom na trvalú zatienenosť a permanentne vysoký stav vody sú cenózy ripikolných a aquikolných nosáčikov druhovo chudobné a málo diverzifikované. Z kvantitatívnej stránky prevládajú charakteristické druhy nad sprievodnými. K charakteristickým patrí druh bahnitých brehov stojatých a pomaly tečúcich vôd (Tanysphyrus lemnae) a dva druhy žijúce na koreniacej pobrežnej vegetácii (Pelenomus waltoni, P. sisymbrii). V menšej miere (najmä po kvantitatívnej stránke) sú zastúpené druhy z okolitého lužného lesa.
Na Kráľovskej lúke bol eudominantom silno hygrofilný druh T. lemnae. Počas päťročného monitoringu sa na tejto lokalite zistila prítomnosť 38 druhov (Tab. 5.7), z toho iba P. sisymbrii, Mononychus punctumalbum a Ceutorhynchus gerhardti boli zastúpené vo vzorkách zo všetkých rokov. Cenóza nosáčikov monitorovacej plochy je vo vývoji, čoho dôkazom je malý počet spoločných druhov zistených počas všetkých rokov monitoringu. Prítomnosť charakteristických druhov (cenobiontov) však indikuje prírodné pobrežné spoločenstvo stojatých vôd. Monitorovaciu plochu je možné z tohto hľadiska hodnotiť ako zachovalú s bohatým zastúpením hygrofilných paludikolných druhov.
Zbery z Istragova sú z pobrežia vysýchajúceho slepého ramena Dunaja, ktoré je v lužnom lese (Salici - Populetum). Vegetácia je zastúpená asociáciou Phragmitetum communis. Väčšie množstvo vody bolo v ramene iba v jarnom období (apríl a máj). Eudominantným bol ubikvista, monofág na pŕhľave Nedyus quadrimaculatus. Z charakteristických druhov tu boli zistené hygrofilné a paludikolné nosáčiky Pelenomus canaliculatus, Bagous collignensis, Bagous subcarinatus, T. lemnae a Phytobius leucogaster. Počas päťročného výskumu sa na monitorovacej ploche v Istragove sa zistilo 39 druhov. Ani jeden sa nevyskytoval počas všetkých piatich rokov. Po roku 1994 sa počet determinovaných druhov stabilizoval (1993 iba 9, 1994 už 17, 1995 – 21, 1996 – 18 a 1997 – 21 druhov). Lokalitu hodnotíme ako silne narušenú. Ak sa zabezpečí prívod vody do tunajšej ramennej sústavy, môže sa viac-menej pôvodné spoločenstvo nosáčikov v pomerne krátkom čase obnoviť, ako vidieť aj z pomerne vyrovnaného zastúpenia charakteristických a sprievodných druhov na jednej strane, ako aj druhov hygrofilných a druhov bez vlhkostnej preferencie. Taxocenózy sú však po kvantitatívnej stránke chudobné a vyznačujú sa najnižšou priemernou abundanciou počas celého obdobia monitoringu ako aj v jednotlivých sezónach (Tab. 5.7).
Tab. 5.7 Celkový prehľad zistených druhov nosáčikov (Coleoptera, Curculionoidae) vo vegetačnom leme dunajských ramien v r. 1993-1997
General review of Coleoptera and Curculionoidae in vegetation edge of the Danube arms in 1993-1997
| Lokality: | 6) Dobrohošť - Dunajské kriviny, | 9) Bodíky, | 10) Kráľovská lúka, |
| 14) Istragov, | 18) Kľúčovec - Sporná sihoť, | 23) Číčov - Starý les; |
| Vysvetlivky: | |||
| Fi – fidelita: | Ch –charakteristický druh, | Sp – sprievodný druh; | |
| To – topická väzba k habitatu: | Ub-ubikvist, | Eu – eurytop, | St – stenotop; |
| Vl – preferencia vlhkosti: | Hy – hygrofil, | Xe – xerofil, | Ne – bez preferencie; |
| Tr –trofická špecializácia imág: | S1-monofág, | S2-užší oligofág, | S3-širší oligofág, |
| G-polyfág |
|
Čeľaď Druh |
6 |
9 |
10 |
14 |
18 |
23 |
Fi |
To |
Vl |
Tr |
|
Attelabidae |
|
|
|
|
|
|
|
|
|
|
|
Pselaphorhynchites nanus (Paykull,1792) |
|
n |
|
|
|
|
Ch |
Eu |
Hy |
G |
|
Pselaphorhynchites tomentosus (Gyllenhal, 1839) |
|
|
|
n |
|
|
Ch |
Eu |
Hy |
S3 |
|
Apionidae |
|
|
|
|
|
|
|
|
|
|
|
Apion brevirostre (Herbst, 1797) |
n |
|
|
|
|
|
Sp |
Eu |
Xe |
S2 |
|
Apion frumentarium (Linnaeus, 1758) |
n |
|
|
|
|
|
Sp |
Eu |
Ne |
S2 |
|
Apion minimum (Herbst, 1797) |
n |
|
|
|
|
|
Ch |
Eu |
Hy |
S2 |
|
Apion urticarium (Herbst, 1784) |
n |
n |
|
|
|
Sp |
Eu |
Xe |
S1 |
|
|
Apion confluens (Kirby, 1808) |
|
|
n |
|
|
|
Sp |
Eu |
Xe |
S3 |
|
Apion onopordi (Kirby, 1808) |
n |
|
n |
n |
|
|
Sp |
Ub |
Ne |
S3 |
|
Apion hookeri (Kirby, 1808) |
|
|
|
|
n |
|
Sp |
Eu |
Xe |
S3 |
|
Apion tenue (Kirby, 1808) |
n |
|
|
|
|
n |
Sp |
Eu |
Xe |
S2 |
|
Apion pisi (Fabricius, 1801) |
n |
|
n |
|
|
Sp |
Eu |
Xe |
S2 |
|
|
Apion punctirostre (Gyllenhal, 1839) |
n |
|
|
|
|
|
Sp |
St |
Xe |
S2 |
|
Apion apricans (Herbst, 1797) |
n |
|
|
|
|
|
Sp |
Eu |
Ne |
S1 |
|
Apion assimile (Kirby, 1808) |
n |
|
|
|
|
n |
Sp |
Ub |
Ne |
S2 |
|
Apion trifolii (Linnaeus, 1768) |
n |
n |
|
|
n |
|
Sp |
Eu |
Xe |
S2 |
|
Curculionidae |
|
|
|
|
|
|
|
|
|
|
|
Otiorhynchus inflatus (Gyllenhal, 1834) |
n |
|
|
|
|
|
Sp |
Eu |
Ne |
G |
|
Phyllobius sinuatus (Fabricius, 1801) |
n |
n |
n |
n |
n |
n |
Ch |
St |
Hy |
S3 |
|
Phyllobius cloropus (Linnaeus, 1758) |
|
|
|
|
n |
|
Sp |
Eu |
Xe |
G |
|
Phyllobius viridearis (Laicharting, 1781) |
|
|
|
n |
|
|
Sp |
Eu |
Ne |
G |
|
Phyllobius oblongus (Linnaeus, 1758) |
n |
n |
|
n |
n |
|
Sp |
Eu |
Ne |
G |
|
Phyllobius calcaratus (Fabricius, 1792) |
|
|
|
n |
|
n |
Ch |
Eu |
Hy |
G |
|
Phyllobius maculicornis (Germar, 1824) |
n |
|
n |
|
|
|
Sp |
Eu |
Ne |
G |
|
Phyllobius pomaceus (Gyllenhal, 1834) |
n |
|
n |
n |
n |
|
Ch |
Eu |
Hy |
S1 |
|
Polydrusus pterygomalis (Boheman, 1840) |
n |
n |
|
n |
|
|
Sp |
Eu |
Ne |
G |
|
Polydrusus sericeus (Schaller, 1783) |
n |
n |
n |
n |
n |
|
Sp |
Eu |
Ne |
G |
|
Eusomus ovulum (Germar, 1824) |
n |
|
|
n |
|
|
Sp |
Eu |
Xe |
G |
|
Sitona hispidulus (Fabricius, 1776) |
|
|
|
|
n |
|
Sp |
Eu |
Ne |
S3 |
|
Sitona humeralis (Stephens, 1831) |
n |
n |
|
|
n |
n |
Sp |
Eu |
Ne |
S3 |
|
Sitona lineatus (Linnaeus, 1758) |
n |
n |
n |
n |
n |
n |
Sp |
Eu |
Ne |
S3 |
|
Sitona macularis (Marsham, 1802) |
n |
n |
n |
n |
n |
n |
Sp |
Eu |
Xe |
S3 |
|
Sitona puncticollis (Stephens, 1831) |
|
|
|
|
|
n |
Sp |
St |
Ne |
S3 |
|
Sitona suturalis (Stephens, 1831) |
n |
n |
n |
n |
n |
n |
Sp |
Eu |
Ne |
S2 |
|
Chlorophanus graminicola (Schönherr, 1832) |
n |
n |
n |
|
|
|
Ch |
St |
Hy |
G |
|
Chlorophanus viridis (Linnaeus, 1758) |
n |
n |
|
|
|
|
Ch |
Eu |
Ne |
G |
|
Tanymecus dilaticollis (Gyllenhal, 1834) |
|
|
|
|
n |
|
Ch |
St |
Ne |
G |
|
Tanymecus palliatus (Fabricius, 1787) |
n |
n |
|
n |
n |
n |
Sp |
Eu |
Ne |
G |
|
Lixus albomarginatus (Boheman, 1843) |
n |
|
|
|
|
|
Sp |
St |
Xe |
S3 |
|
Lixus bardanae (Fabricius, 1787) |
|
|
n |
|
|
n |
Ch |
St |
Hy |
S2 |
|
Lixus fasciculatus (Boheman, 1836) |
n |
|
|
|
|
|
Sp |
St |
Ne |
S3 |
|
Lixus paraplecticus (Linnaeus, 1758) |
|
|
|
|
n |
|
Ch |
St |
Hy |
S3 |
|
Larinodontes sturnus (Schaller, 1783) |
n |
|
|
|
|
|
Sp |
St |
Ne |
S3 |
|
Larinodontes turbinatus (Gyllenhal, 1836) |
n |
|
|
|
|
|
Sp |
St |
Xe |
S3 |
|
Cossonus linearis (Fabricius, 1775) |
|
|
|
n |
|
|
Ch |
Eu |
Hy |
G |
|
Dicranthus majzlani (Kodada, Holecová et Behne, 1992) |
|
|
|
|
|
n |
Ch |
St |
Hy |
S1 |
|
Bagous angustus (Silfverberg, 1977) |
|
|
n |
|
|
|
Ch |
St |
Hy |
S2 |
|
Bagous argillaceus (Gyllenhal, 1836) |
|
|
n |
|
|
n |
Ch |
St |
Hy |
S2 |
|
Bagous collignensis (Herbst, 1797) |
|
|
n |
n |
n |
n |
Ch |
St |
Hy |
S2 |
|
Bagous limosus (Gyllenhal, 1827) |
|
|
|
|
n |
n |
Ch |
St |
Hy |
S2 |
|
Bagous subcarinatus (Gyllenhal, 1836) |
|
n |
n |
|
n |
n |
Ch |
St |
Hy |
S1 |
|
Bagous tempestivus (Herbst, 1795) |
|
|
n |
|
n |
n |
Ch |
Eu |
Hy |
S3 |
|
Bagous bagdatensis (Pic, 1904) |
|
|
|
|
|
n |
Ch |
St |
Hy |
S2 |
|
Bagous glabrirostris (Herbst, 1795) |
|
|
|
|
|
n |
Ch |
Eu |
Hy |
S3 |
|
Tanysphyrus lemnae (Paykull, 1792) |
|
n |
n |
n |
n |
n |
Ch |
St |
Hy |
S2 |
|
Dorytomus affinis (Paykull, 1800) |
|
|
n |
|
|
|
Ch |
St |
Hy |
S2 |
|
Dorytomus longimanus (Forster, 1771) |
|
|
|
n |
|
|
Ch |
Eu |
Hy |
S2 |
|
Dorytomus ictor (Herbst, 1795) |
|
n |
|
n |
|
|
Ch |
Eu |
Hy |
S2 |
|
Dorytomus hirtipennis (Bedel, 1884) |
|
|
|
|
n |
|
Ch |
St |
Hy |
S2 |
|
Dorytomus tremulae (Fabricius, 1787) |
|
n |
n |
|
|
|
Ch |
St |
Hy |
S2 |
|
Dorytomus melanophthalmus (Paykull, 1792) |
n |
n |
n |
|
|
|
Ch |
Eu |
Hy |
S2 |
|
Notaris bimaculatus (Fabricius, 1787) |
|
|
|
|
n |
n |
Ch |
St |
Hy |
S2 |
|
Notaris scirpi (Fabricius, 1792) |
|
|
|
|
|
n |
Ch |
St |
Hy |
S3 |
|
Grypus equiseti (Fabricius, 1775) |
n |
|
n |
|
|
n |
Ch |
Eu |
Hy |
S2 |
|
Smicronyx coecus (Reich, 1797) |
|
n |
|
|
|
|
Sp |
St |
Xe |
S2 |
|
Smicronyx jungermanniae (Reich, 1797) |
|
|
|
n |
|
|
Sp |
St |
Xe |
S2 |
|
Tychius quinquepunctatus (Linnaeus, 1758) |
|
|
|
|
|
n |
Sp |
Eu |
Xe |
S3 |
|
Tychius meliloti (Stephens, 1831) |
|
|
|
|
|
n |
Sp |
St |
Xe |
S2 |
|
Tychius picirostris (Fabricius, 1787) |
|
|
|
n |
|
n |
Sp |
Eu |
Ne |
S2 |
|
Tychius stephensi (Schönherr, 1836) |
n |
|
|
|
|
|
Sp |
St |
Xe |
S2 |
|
Anthonomus rubi (Herbst, 1795) |
n |
|
|
|
|
|
Sp |
Ub |
Ne |
S3 |
|
Curculio crux (Fabricius, 1776) |
|
n |
|
|
|
|
Ch |
Eu |
Hy |
S2 |
|
Curculio salicivorus (Paykull, 1792) |
|
n |
|
n |
|
|
Ch |
Eu |
Hy |
S2 |
|
Lepyrus armatus (Weise, 1893) |
n |
|
|
n |
|
|
Ch |
St |
Hy |
S3 |
|
Lepyrus palustris (Scopoli, 1763) |
|
n |
|
|
|
|
Ch |
St |
Hy |
S2 |
|
Hylobius transversovittatus (Goeze, 1771) |
|
|
|
|
n |
|
Ch |
St |
Hy |
S1 |
|
Alophus weberi (Penecke, 1901) |
|
|
|
n |
|
|
Ch |
St |
Hy |
S3 |
|
Hypera postica (Gyllenhal, 1813) |
|
|
|
|
n |
|
Sp |
Eu |
Ne |
S3 |
|
Hypera rumicis (Linnaeus, 1758) |
n |
n |
n |
|
|
|
Ch |
Eu |
Hy |
S2 |
|
Baris coerulescens (Scopoli, 1763) |
n |
|
|
n |
|
|
Sp |
Eu |
Ne |
S3 |
|
Limnobaris dolorosa (Goeze, 1777) |
|
|
n |
|
|
|
Ch |
St |
Hy |
S3 |
|
Mononychus punctumalbum (Herbst, 1784) |
|
|
n |
|
|
n |
Ch |
St |
Hy |
S2 |
|
Phytobius leucogaster (Marsham, 1805) |
|
|
n |
n |
|
n |
Ch |
St |
Hy |
S2 |
|
Pelenomus canaliculatus (Fahraeus, 1846) |
|
|
n |
n |
n |
n |
Ch |
St |
Hy |
S2 |
|
Pelenomus waltoni (Boheman, 1843) |
|
n |
|
|
|
|
Ch |
St |
Hy |
S2 |
|
Rhinoncus albicinctus (Gyllenhal, 1837) |
n |
|
n |
|
n |
n |
Ch |
St |
Hy |
S1 |
|
Rhinoncus bosnicus (Schultze, 1900) |
|
|
n |
n |
n |
n |
Ch |
St |
Hy |
S2 |
|
Rhinoncus inconspectus (Herbst, 1795) |
n |
n |
|
|
n |
n |
Ch |
St |
Hy |
S1 |
|
Rhinoncus pericarpius (Linneaus, 1758) |
n |
n |
n |
n |
n |
n |
Sp |
Eu |
Ne |
S2 |
|
Rhinoncus perpendicularis (Reich, 1797) |
|
|
n |
n |
n |
n |
Ch |
Eu |
Ne |
S2 |
|
Poophagus sisymbrii (Fabricius, 1777) |
n |
n |
n |
n |
n |
n |
Ch |
St |
Hy |
S2 |
|
Tapinotus sellatus (Fabricius, 1794) |
|
|
|
|
n |
Ch |
St |
Hy |
S1 |
|
|
Ceutorhynchus contractus (Marsham, 1802) |
|
n |
|
|
|
Sp |
Ub |
Ne |
G |
|
|
Ceutorhynchus erysimi (Fabricius, 1787) |
|
|
n |
n |
Sp |
Ub |
Ne |
S3 |
||
|
Ceutorhynchus floralis (Paykull, 1792) |
|
n |
|
n |
Sp |
Ub |
Ne |
S3 |
||
|
Ceutorhynchus gerhardti (Schultze, 1899) |
n |
n |
n |
n |
n |
n |
Sp |
St |
Xe |
S1 |
|
Ceutorhynchus obstrictus (Marsham, 1802) |
n |
|
|
|
|
n |
Sp |
Ub |
Ne |
S3 |
|
Ceutorhynchus pallidactylus (Marsham, 1802) |
n |
n |
n |
n |
n |
n |
Sp |
Ub |
Ne |
S3 |
|
Ceutorhynchus rapae (Gyllenhal, 1837) |
|
|
|
|
n |
Sp |
Eu |
Ne |
S3 |
|
|
Ceutorhynchus roberti (Gyllenhal, 1837) |
n |
|
|
|
|
|
Sp |
St |
Ne |
S1 |
|
Ceutorhynchus syrites (Germar, 1824) |
|
|
|
|
n |
Sp |
St |
Xe |
S3 |
|
|
Parethelcus pollinarius (Forster,1771) |
n |
|
n |
|
|
Ch |
St |
Hy |
S1 |
|
|
Mogulones symphyti (Bedel, 1885) |
|
|
n |
n |
|
Ch |
St |
Hy |
S1 |
|
|
Nedyus quadrimaculatus (Linnaeus, 1758) |
n |
n |
n |
n |
n |
Sp |
Ub |
Ne |
S1 |
|
|
Nanophyes brevis (Boheman, 1845) |
|
|
|
|
|
n |
Ch |
St |
Hy |
S1 |
|
Nanophyes globiformis (Kiesenwetter, 1864) |
|
|
|
|
n |
n |
Ch |
St |
Hy |
S2 |
|
Nanophyes marmoratus (Goeze, 1777) |
n |
|
n |
n |
n |
n |
Ch |
St |
Hy |
S2 |
|
Gymnetron pascuorum (Gyllenhal, 1813) |
|
|
|
|
n |
Sp |
St |
Xe |
S1 |
|
|
Gymnetron veronicae (Germar, 1821) |
|
|
|
n |
n |
Ch |
St |
Hy |
S2 |
|
|
Gymnetron villosulum Gyllenhal, 1837 |
|
|
|
|
n |
Ch |
St |
Hy |
S2 |
|
|
Gymnetron asellus (Gravenhorst, 1807) |
n |
|
|
|
|
Sp |
St |
Xe |
S2 |
|
|
Gymnetron bipustulatum (Rossi, 1792) |
n |
|
|
|
|
Sp |
St |
Xe |
S2 |
|
|
Gymnetron tetrum (Fabricius, 1801) |
n |
|
|
|
|
Sp |
St |
Xe |
S2 |
|
|
Cionus olivieri (Rosenschoeld, 1838) |
n |
|
|
|
|
Sp |
St |
Xe |
S1 |
|
|
Rhamphus pulicarius (Herbst, 1795) |
|
|
|
n |
|
Ch |
Eu |
Ne |
S3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Počet zistených druhov |
51 |
32 |
38 |
39 |
38 |
49 |
113 |
|
|
|
|
Počet zozbieraných jedincov |
872 |
3447 |
608 |
222 |
1008 |
1215 |
7372 |
|
|
|
|
Priemerná abundancia |
33,53 |
164,14 |
24,32 |
9,65 |
42 |
48,60 |
|
|
|
|
|
|
||||||||||
|
Fidelita |
||||||||||
|
Počet charakteristických druhov |
13 |
17 |
24 |
21 |
22 |
29 |
|
|
|
|
|
Počet sprievodných druhov |
38 |
15 |
14 |
18 |
16 |
20 |
|
|
|
|
|
Topická väzba k habitatu |
||||||||||
|
Počet ubikvistických druhov |
6 |
2 |
5 |
4 |
2 |
5 |
|
|
|
|
|
Počet eurytopných druhov |
26 |
18 |
14 |
21 |
16 |
15 |
|
|
|
|
|
Počet stenotopných druhov |
19 |
12 |
19 |
14 |
20 |
29 |
|
|
|
|
|
Preferencia vlhkosti |
||||||||||
|
Počet hygrofilných druhov |
12 |
16 |
23 |
19 |
20 |
28 |
|
|
|
|
|
Počet xerofilných druhov |
16 |
5 |
4 |
4 |
5 |
7 |
|
|
|
|
|
Počet druhov bez vlhkostnej preferencie |
23 |
11 |
11 |
16 |
13 |
14 |
|
|
|
|
|
Trofická špecializácia |
||||||||||
|
S1 – počet druhov |
10 |
6 |
5 |
5 |
8 |
13 |
|
|
|
|
|
S2 - počet druhov |
18 |
14 |
19 |
15 |
14 |
26 |
|
|
|
|
|
S3 - počet druhov |
14 |
5 |
10 |
11 |
11 |
17 |
|
|
|
|
|
G - počet druhov |
9 |
7 |
4 |
8 |
5 |
3 |
|
|
|
|
Podľa výsledkov monitoringu nosáčikov v období po prehradení Dunaja môžeme monitorovacie plochy rozdeliť na tri skupiny:
1. prírodne zachovalé, s bohatými taxocenózami fytofágnych Coleoptera a so značným percentuálnym zastúpením hygrofilných druhov (vrátane semiakvatických a akvatických), z ktorých mnohé sú indikátormi prírodne zachovalých stojatých a mierne tečúcich nížinných vôd (napr. rody Bagous, Tanysphyrus, Limnobaris, Notaris, Poophagus, Phytobius, Rhinoncus, Pelenomus). Do tejto skupiny môžeme zaradiť ramená v Bodíkoch – Bodíckej bráne a referenčné lokality mimo vplyvu VD Gabčíkovo (Kľúčovec - Sporná sihoť a Čičov - Starý les).
2. plochy s nestabilizovanými taxocenózami nosáčikov, ktoré sú po prehradení Dunaja v sukcesii. Do tejto kategórie patrí rameno na lokalite Bodíky-Kráľovská lúka, kde sme pozorovali pozitívny vývoj v súvislosti so zlepšujúcimi sa pomermi vplyvom simulovaných záplav.
3. silne ruderalizované a aridizované, do tejto skupiny patria vyschnuté rameno v Dobrohošti-Dunajských krivinách a periodicky vysýchajúce lesné ramená na Istragove. Prevládajú tu druhy xerofilné, ruderálne a v menšej miere aj druhy z okolitých lužných lesov. Najmarkantnejšie sa to prejavuje na lokalite v Dobrohošti, kde došlo k prudkému poklesu počtu druhov. Úplne absentuje druh P. sisymbrii troficky viazaný na Rorippa amphibia, aj napriek skutočnosti, že hostiteľská rastlina sa na lokalite ešte stále vyskytuje. Ešte v r. 1993 a 1994 tento druh na lokalite dosahoval pomerne vysoké hodnoty dominancie, od r. 1995 sa na lokalite prestal vyskytovať.
Ornitocenózy záujmového územia sa pôvodne vyznačovali vysokou diverzitou a denzitou druhov. Hniezdnu ornitocenózu podunajských lužných lesov pred výstavbou VD Gabčíkovo tvorilo 104 druhov vtákov, medzi nimi viacero významných hniezdičov (Milvus migrans, Aythya nyroca, Ixobrychus minutus, Ciconia nigra, Pernis apivorus, Alcedo atthis, Dendrocopos medius). Zmeny, ktoré v ornitocenóze podunajských lužných lesov a celej vnútrozemskej delte Dunaja nastali výstavbou VD Gabčíkovo, boli podmienené zánikom časti ramennej sústavy a následne (na jej mieste) vznikom nového biotopu Čunovskej zdrže, ako aj adaptívnymi zmenami ekosystému na nové hydrologické pomery v zachovalej ramennej sústave medzi Dobrohošťou a Sapom. Ďalej je treba zdôrazniť, že na zmeny, ktoré v ornitocenóze podunajských lužných lesov prebiehajú, má zásadný vplyv aj lesné hospodárstvo spôsobom ťažby.
Výstavbou Čunovskej (Hrušovskej) zdrže zanikol komplex lužných lesov, dunajských ostrovov a ramien pri Podunajských Biskupiciach, Hamuliakove, Šamoríne a Šamoríne – Čilistove. Hniezdnu ornitocenózu týchto lesov pred odlesnením tvorilo 77 druhov vtákov, z významnejších druhov napr. haja tmavá (Milvus migrans) sokol lastovičiar (Falco subbuteo) ďateľ prostredný (Dendrocopos medius), penica jarabá (Sylvia nisoria). Výstavbou gabčíkovskej elektrárne a odpadového kanála zanikli zasa pri Gabčíkove mŕtve ramená Dunaja, ktoré z hľadiska zoologického významu presahovali rámec regiónu. Hniezdnu ornitocenózu týchto lokalít pred ich zánikom tvorilo 20 druhov vtákov, z ktorých niektoré predstavovali v našich podmienkach, z hľadiska ochrany genofondu, druhy celoslovenského významu, napr. chriašteľ malý (Porzana parva), chochlačka bielooká (Aythya nyroca).
Po prehradení koryta Dunaja v októbri 1992, vznikom Čunovskej zdrže vznikol nový biotop – vodná plocha o rozlohe 38 km2. Už v prvých rokoch existencie tohoto nového biotopu sa zdrž ukázala ako lokalita významná pre ťah a zimovanie vodných vtákov. Vtáky preťahujúce Dunajom atrahuje k dlhšej zastávke a väčšej koncentrácii rozľahlá vodná plocha. Na zdrži sa počas ťahu sústreďujú kŕdle kormoránov, husí, kačíc, potápačov, čajok, bahniakov. Mnohé druhy vodných vtákov tu zimujú. Pre hniezdenie vtákov bol uprostred zdrže vytvorený umelý ostrov (5,7 ha veľká depónia), kde vznikla kolónia čajky smejivej (Larus ridibundus) a rybára riečneho (Sterna hirundo). Na Hrušovskej zdrži zahniezdili aj ďalšie expanzívne druhy ako chochlačka vrkočatá (Aythya fuligula), hrdzavka potápavá (Netta rufina), čajka bielohlavá (Larus cachinnans), čajka čiernohlavá (Larus melanocephalus), podobný proces v súčasnosti prebieha aj na iných lokalitách na Slovensku (napr. Sĺňava pri Piešťanoch).
Po uvedení VD Gabčíkovo do prevádzky došlo v území z hľadiska močiarnych biotopov, významných pre hniezdenie vtákov, k prestúpeniu lokalít. Niektoré lokality (napr. ostrov Istragov pri Gabčíkove) vyschli, na odstavenej ramennej sústave, po rozrastení sa trsťových porastov však vznikli nové (napr. močiar Forrás pri Bodíkoch).
Lokalita Istragov plnila významnú úlohu loviska pre volavkovité vtáky (predovšetkým pre druhy beluša veľká (Egretta alba), volavka popolavá (Ardea cinerea) a bocian čierny (Ciconia nigra) v čase po vyletení mláďat a pred migráciou. Po nízkej hladine vody vtáky využívali rozsiahle terénne depresie so zvyškovou plytkou vodou na uľahčený lov potravy. Tento jav sa vyskytoval aj v ostatných častiach ramennej sústavy, na Istragove však bol najčastejší. V auguste a septembri tu neraz lovilo do 200 beluší veľkých a niekoľko desiatok (až 70 – 80) bocianov čiernych. Po vylúčení dynamiky vodného režimu tento jav z tohto úseku Dunaja vymizol. Lokalita Forrás plní v súčasnosti funkciu hniezdiska pre hniezdiče v litorálnej vegetácii, predovšetkým trsti (Bohuš 1999).
Po uvedení VD do prevádzky vymizli z ramenného systému ako hniezdiče dva druhy vtákov – brehuľa hnedá (Riparia riparia) a kalužiak riečny (Actitis hypoleucos). Brehuľa hnedá tu hniezdila v brehoch dunajských ramien. Po uvedení Vodného diela došlo k úplnému zániku aj miernej laterálnej erózie, brehy ramien zarástli a stali sa pre hniezdenie brehule nevhodné. Podobne hniezdne biotopy kalužiaka riečneho, ktorý tu hniezdil na štrkopieskových laviciach a brehoch, zarástli a stali sa pre hniezdenie tohoto druhu nevhodné.
K negatívnym zmenám v oblasti podunajských lužných lesov dochádza v poslednom období nadmernou ťažbou lesných porastov (holoruby), čím zanikajú hniezdne možnosti pre dravce, bociany čierne a dutinové hniezdiče. Pri niektorých druhoch tak dochádza k výraznému poklesu početnosti – napr. v 80. rokoch hniezdilo na slovenskej strane podunajských lužných lesov 13 – 15 párov bocianov čiernych, v roku 2001 tu hniezdili len tri páry!
Aj napriek zmenám, ktoré v ornitocenóze podunajských lužných lesov nastali, predstavuje celá oblasť ramenného systému s lužnými lesmi a mozaikou močiarnych biotopov, z ornitologického hľadiska, aj v súčasnosti územie s nadregionálnym významom. Je však žiadúce pristúpiť k jeho renaturácii.
Fauna cicavcov (Mammalia) sa po výstavbe VDG druhovým zložením podstatne nezmenila, nastali však zmeny v jej štruktúre. Taxocenóza drobných zemných cicavcov je tvorená druhmi Sorex araneus, Apodemus flavicollis, Clethrionomys glareolus, Sorex minutus, Crocidura leucodon, Crocidura suaveolens, Microtus arvalis, Microtus oeconomus, Pitymys subterraneus, Apodemus sylvaticus, Micromys minutus, antropogénne je podmienené prenikanie nepôvodných M. arvalis a M. musculus.
Jadrom taxocenózy drobných zemných cicavcov sú štyri druhy: A. flavicollis, S. araneus, C. glareolus a A. sylvaticus s výraznou dominanciou prvých troch univerzálnych lesných druhov. Rozhodujúci vplyv na štruktúru dominancie tejto synúzie majú záplavy resp. ich absencia, v závislosti najmä na vlhkostnom gradiente sa mení štruktúra dominancie. Najlepšie prežíva záplavy ryšavka žltohrdlá (A. flavicollis), po záplave rýchle regenerujú i populácie S. araneus a C. glareolus. Pri absencii záplav absolútnu dominanciu preberá druh C. glareolus, ktorého záplavy tlmia. Do pozadia ustupuje nielen S. araneus, ale aj A. flavicollis. Pri poklese hladiny podzemnej vody klesá úživnosť biotopu a s tým súvisí znížená celková početnosť a biomasa mikromammálií.
Je zrejmé, že synúzie v ramennej sústave prežívajú dlhodobý stres prejavujúci sa úplne neprirodzenou dominanciou jedného druhu C. glareolus. Počas prvých desiatich rokov po úpravách v ramennej sústave C. glareolus sa výrazne rozmnožil na úkor A. flavicollis, ktorý v minulosti spolu so S. araneus a C. glareolus tvorili 80 až 90% biomasy mikromammalií. Pred sprevádzkovaním vodného diela záplavy znižovali početnosť C. glareolus, pretože jeho schopnosť zachrániť sa vylezením na dreviny je oveľa nižšia ako u A. flavicollis, ktorý v minulosti početne prevládal. Nežiadúcosť takéhoto stavu je aj v tom, že C. glareolus je závažným konkurentom v priestore aj v potrave reliktnému a kriticky ohrozenému druhu M. oeconomus, ktorého početný stav po sprevádzkovaní vodného diela poklesol tak kriticky, že do popredia vystupuje otázka možnosti jeho prežitia na území Slovenska. A. flavicollis takým závažným konkurentom pre M. oeconomus nebol, pretože sa živí prevažne semenami. Preto sa v minulosti M. oeconomus, i keď nikdy nie hojne, vyskytoval na všetkých vhodných stanovištiach v oblasti ramennej sústavy a aj po záplavách sa jeho populácia veľmi rýchlo zregenerovala.
V situácii, keď záplavy zdvihnú na krátky čas hladinu o 30-40 cm (a neplnia iné funkcie, najmä zásobovanie živinami), môžu mať v nevhodnom čase aj zhubný efekt práve na najcitlivejšie a najvzácnejšie elementy teriofauny, pre ktoré je vhodnejšie udržovať celoročne ustálenú hladinu vôd. Záplavy nemajú na ne negatívny vplyv v jesennom období (v tomto období sú záplavy netypické), kedy už nie sú ohrozené hniezda s mláďatami reliktného a kriticky ohrozeného druhu M. oeconomus. Jeho početnosť na vhodných lokalitách v ramennej sústave výrazne klesla a na väčšine svojich bývalých lokalít sa dnes už nevyskytuje. Prvotnou príčinou bol fakt, že v zime 1992-93 celá ramenná sústava vyschla a posledné mláčky so stojatou vodou premrzli až do dna. Takisto premrzla aj porézna piesčitá pôda s nehlbokými hniezdami drobných cicavcov, predovšetkým M. oeconomus. Hlboké nory si nikdy v týchto miestach vyrývať nemohli pre obvykle vysokú hladinu podzemnej vody, ktorá však udržovala teplotu neklesajúcu pod bod mrazu. Mrazy a eliminácia podzemnej vody mali za následok, že hniezda v nehlbokých norách na okrajoch vodných nádrží premrzli aj s ich obyvateľmi. Konkurenčné druhy, Apodemus flavicollis, ale hlavne Clethrionomys glareolus týmto neboli postihnuté, pretože prevažná väčšina ich populácie mala hniezdne nory v suchších, vyššie situovaných polohách a zimu bez problémov prežili. Počas jari a v lete sa dobre prezimovavší a primerane rozmnožený C. glareolus začal šíriť aj do miest obývaných pôvodne hrabošom severským a vytláčal odtiaľ posledné jedince. Naviac nenasledovala už žiadna taká záplava, ktorú by M. oeconomus prežil úspešnejšie, ako C. glareolus. Tretím faktorom znižujúcim početnosť už zdecimovaných populácií M. oeconomus bolo vykonanie umelej záplavy v jarných mesiacoch. Vzdutie hladiny o 40 cm postihlo iba druhy, resp. jedince vychovávajúce potomstvo tesne pri vode, najviac M. oeconomus.
Ďalšou zmenou je, že v súčasnosti takmer absentuje v ramennej sústave tretí najpočetnejší druh lužných lesov – S. araneus.
V osemdesiatych rokoch minulého storočia sa významnou súčasťou teriofauny stal typický semiakvatický cicavec bobor (Castor fiber), ktorý môže zaujímať význačné postavenie pri formovaní bioty lužných lesov. Na Slovensko prenikol pri reštitúcii v rakúskom Eckartsau v roku 1987. Šíril sa po prúde Dunaja. Niektoré zvieratá prenikali do rieky Moravy a z nej do jej prítokov z Borskej nížiny, iné migrovali ďalej dole po prúde a vstupovali do dunajských ramien. . Zatial čo bobry na Záhorskej nížine našli dobré podmienky pre prežitie, vytvorili páry a začali sa úspešne rozmnožovať, bobry v dunajských ramenách boli po usadení odnesené najbližšou povodňou. Pomery v dunajských ramenách sa zmenili po roku 1992 , kedy bolo dokončené VDG a staré koryto Dunaja bolo pri Čunove prehradené. Ramenná sústava predstavuje v súčasnom stave pre bobry ideálny biotop. Čunovská zdrž predstavuje však pre bobry dodnes neprechodnú prekážku, a tak, zatiaľ čo počet bobrov na Borskej nížine z roka na rok narastal, ideálne bobrie habitaty v ramennej sústave ostávali pre bobry neprístupné. V súčasnosti (rok 2002) registrujeme v ramennej sústave iba 3-4 obývané bobrie revíry, z ktorých o jednom vieme, že je obývaný reprodukujúcim sa párom. Predpokladáme, že tieto zvieratá prenikli do ramennej sústavy obchvatom – prechodom cez hrebeň Malých Karpát a odtiaľ stekajúcimi potokmi až do Dudváhu a ďalej cez Čiernu Vodu a Vážsky Dunaj až do hlavného toku Dunaja a odtiaľ proti prúdu až do ramennej sústavy (dva obývané revíry boli zistené aj na Malom Dunaji).
Súhrnne možno povedať, že najväčšie zmeny v terestrickej faune boli zistené na monitorovacej ploche Dunajské kriviny pri Dobrohošti (Foto 9 až po Foto 23) na úrovni začiatku derivačného kanála. Drénom a zánikom pôvodného hladinového režimu pri absencii záplav prechádza ekosystém vážnymi zmenami. Vzhľadom na nedostatočnú výšku hladiny v najhornejšej časti kanála pod odberným objektom vo vzťahu k prahu stavidla a absencii patričnej manipulácie na tomto objekte, nie sú tu simulované povrchové záplavy, a s výnimkou obdobia riadených záplav nedochádza ani k zavodneniu obvodového ramena. Požadovaný dnový prah nebol vybudovaný. Posledná záplava, ktorá mala vplyv na väčšinu monitorovacej plochy bola v r. 1991, nasledujúca až v roku 2002. Pôvodne navrhovaná pretekaná prehrádzka v Dunaji pri Dunakiliti, ktorá mohla situáciu zlepšiť, bola vybudovaná 2 km vyššie, nad haťou Dunakiliti. Pôvodne bola lokalita pokrytá typickou brestovou jaseninou s topoľom. Vrchné vrstvy pôdneho profilu tvorili hrubé vrstvy naplaveného piesku, pôdna vlhkosť sa pohybovala od 15% – 25%. Po roku 1992 drenážny účinok starého koryta Dunaja viedol k zániku povrchových vôd, poklesu hladiny podzemnej vody a poklesu pôdnej vlhkosti na 0-20%. Pokiaľ ešte v r. 1991 sa HPV pohybovala v hĺbke 2,5 - 3 m, od novembra r. 1992 sa s minimálnymi výkyvmi stabilizovala v hĺbke 4,2 m, čím fakticky nastali podmienky pre vznik tvrdého luhu. Celkove v terestrických spoločenstvách došlo k úbytku hygrofilných a typických lesných druhov a prieniku xenocénov. Mierne zvýšenie pôdnej vlhkosti po simulovanej záplave sa priaznivejšie prejaví najmä na spoločenstvách euedafónu (Acari, Oniscidea, Chilopoda), v menšej miere na epigeických taxocenózach (Coleoptera, Heteroptera). Zlepšenie vlhkostných pomerov sa prejavilo aj na zložení taxocenózy drobných zemných cicavcov. Zo získaných dát sa predbežne nedá usúdiť, či trend zmierňovania aridizácie pozorovaný v posledných rokoch monitoringu je trvalý, alebo sa jedná o fluktuačný výkyv podmienený klímou. V súčasnosti patria terestrické spoločenstvá k najviac degradovaným na celom monitorovanom území. Vykazujú výrazný vplyv antropogénneho faktora a revitalizačný proces postupuje len pomaly.
{kind=link}
{kind=link}
Bodícka brána (Foto 19-28) v nižšie položenej časti derivačného územia je lokalita s brestovou jaseninou s topoľmi a vŕbami, vyšším stupňom pôdnej vlhkosti (25-35%), HPV je v hĺbke 2 – 2,5 m. Po prehradení Dunaja býva monitorovacia plocha sporadicky zaplavovaná. Výrub porastu šľachteného topoľa viedol k nežiaducemu presvetleniu. Dôsledkom sú zmeny v skladbe bylinnej aj krovinovej etáže vegetácie, na ktoré nadviazali zmeny v zložení spoločenstiev terestrických živočíchov – prienik druhov otvorených priestranstiev, pokles počtu hygrofilných druhov, zvyšovanie zastúpenia mezohygrofilných a euryekných druhov; mnohé z hygrofilných druhov sú na hranici sledovateľnosti (Mollusca, Coleoptera). Tento pokles početnosti sa pri časti druhov spomalil až zvrátil a u niektorých hygrofilov sa v poslednom roku monitoringu abundancia zvýšila (Coleoptera, Lepidoptera, Oniscidea, Chilopoda). Vysokú dominanciu si zachovávajú euryvalentné a vlhkostne menej náročné druhy (Lepidoptera). Stav spoločenstiev možno aj napriek uvedeným zmenám ešte stále považovať za reverzibilný (zvýšenie abundancie niektorých vlhkomilných druhov). Zdá sa že sukcesia spoločenstva sa v posledných dvoch rokoch vracia k stavu, ktorý nastal bezprostredne po prevedení hlavného toku Dunaja do derivačného kanála. Trvalosť tohto trendu je však ťažké predpovedať a závisí od interakcie zmeny vlhkosti pôdy, ťažby a výkyvom klimatických faktorov.
Monitorovacia plocha Bodíky – Kráľovská lúka (Foto 56, Foto 57, Foto 58, Foto 59, Foto 60 a Foto 61) je v oblasti derivácie najviac vzdialená od prietočných ramien, bola pravidelne každoročne (niekedy aj dvakrát) zaplavovaná a do roku 1997 závislá od povodňovej vody. Do uvedeného roku bol v suchých rokoch pozorovaný mierny pokles hladiny podzemnej vody a presýchanie pobrežnej močaristej zóny. Plesiopotamal v jadre lokality, oddeľuje zreteľne suchšie stanovište ležiace bližšie k hrádzi a porastané topoľovou monokultúrou od vlhkého mäkkého luhu (Salici-Populetum). Súčasná štruktúra suchozemských spoločenstiev je odrazom hydropedologických pomerov na lokalite za predchádzajúce roky. Zmeny v druhovej skladbe spoločenstva signalizujú neustálenosť podmienok. Celkový trend poklesu druhovej diverzity (Oniscidea, Coleoptera) a vzostupu dominancie mezofilných a nenáročných aj xenocénnych druhov (Mollusca, Coleoptera) však pokračuje, i keď tento pokles sa v niektorých taxocenózach spomalil resp. zastavil (Chilopoda, Coleoptera, Lepidoptera), čo signalizuje zastavenie resp. spomalenie negatívnych trendov sukcesie. Je však zrejmé, že ide o dôsledok dvoch výrazne vlhších vegetačných období, pričom vegetačné obdobie 1997 prinieslo aj dlhodobú záplavu. Celkove dochádza k sukcesii spoločenstiev smerom k spoločenstvám typu suchších mäkkých lužných lesov. V rámci lokality sa v závislosti na zmenách vlhkosti a na drobných rozdieloch v nadmorskej výške (rádovo 50-60 cm) výrazne menilo priestorové rozšírenie druhov. Po vysušení lokality a ústupe silne vlhkomilných druhov zaujali ich miesto v nižšie položených častiach tolerantnejšie druhy, pričom vyvýšené miesta boli výrazne menej osídlené. Zmeny, ktoré nastali v ornitocenóze nemožno pripísať len prevádzkovaniu vodného diela, ale boli ovplyvnené aj výrazne pôsobiacim systémom lesného obhospodarovania.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Lokalita Istragov na konci úseku derivácie patrila do roku 1992 k výrazne hygrickým stanovištiam mäkkého luhu (Salici-Populetum myosotidetosum Jurko, 1958) so skupinami jelší na viacerých miestach. Bližšie k starému korytu Dunaja sú topoľové monokultúry, v ktorých sa v rokoch monitoringu robila intenzívna ťažba. Lokalitu pokrývala vrstva naplaveného piesku. Po prehradení Dunaja a znížení prietoku v jeho pôvodnom koryte došlo k zaklesnutiu hladiny podzemnej vody, k poklesu vlhkosti vo vrchnej vrstve pôdy z 25 – 35% približne o 15%, redukcii plytkých zaplavených depresií, kde nastupuje xeroséria (Lisický 1995, Ševčíková 1997) a sekundárna sukcesia smeruje aj na nižšie položených miestach k spoločenstvu suchšieho variantu mäkkého luhu (Salici-Populetum typicum). K procesu vysychania lokality prispel tiež veľkoplošný výrub topoľov a presvetlenie územia. V terestrických spoločenstvách sa prejavuje ústup druhov indikujúcich podmáčané biotopy resp. druhov vlhkomilnejších (Mollusca, Coleoptera), nahrádzajú ich lesné hygrofilné druhy menej náročné na vlhkosť, čo poukazuje na trend vysúšania stanovišťa. Tieto zmeny súvisia aj s masívnou ťažbou až holorubom a následnými zmenami aj v ostatných etážach. Trend výrazného ústupu silne vlhkomilných druhov a aridizácie sa však v roku 1996 v niektorých skupinách (Mollusca, Lepidoptera) zmiernil a adaptívne sukcesné zmeny sú v porovnaní so zmenami vegetácie pomalšie (Mollusca, Chilopoda). Z hľadiska zastúpenia jednotlivých ekologických skupín sa spoločenstvo na Istragove javí zo sledovaných lokalít ako najmenej postihnuté.
Výstavbu VD sprevádzal rad zmien v prírodnom prostredí, ktoré ovplyvnili prietokový režim Dunaja a jeho ramien a následne sa výrazne odrazili v štruktúre hydrofauny. Došlo k dvom zmenám v prietokovom režime starého koryta Dunaja a to v roku 1992 a v roku 1995. V Dunaji (v úseku Gabčíkovo- Medveďov) došlo ku kolísaniu prietoku (0.5 m výkyv hladiny) v závislosti od prietokov v Bratislave a v závislosti na spúšťaní a zastavovaní prevádzky turbín elektrárne. V Dunaji v úseku nad sútokom s kanálom z elektrárne dochádzalo najmä do roku 1995 k tzv. spätnému vzdutiu hladiny (pri celkovom miernom znížení hladiny) a k zvýšenému usadzovaniu hlavne jemnozrnnejších sedimentov. V hornom úseku Dunaja (Dunajské kriviny - Bodíky) dochádza po roku 1995, v dôsledku regulácie prietokov do maďarskej ramennej sústavy vo vegetačnom období, ku krátkodobému kolísaniu hladín (0,5-0,75 m) v 5 až 10 dňových intervaloch. V slovenskej ramennej sústave došlo po roku 1992 k poklesu hladín, ktorý sa však od mája 1993 eliminoval vodou z derivačného kanála cez nápustný objekt pri Dobrohošti. Rozkolísanosť hladín je pomerne malá s výnimkou riadených záplav a povodní, napríklad v roku 1997. Dochádza tiež k zmene teplotného režimu Dunaja (Dobrohošť-Bodíky) V zimnom období teplota vôd v starom koryte Dunaja neklesá pod 3,5o C ako dôsledok refiltrácie teplejších podzemných vôd, ktoré zo zdrže infiltrovali v letnom období, do menšieho prietoku v starom koryte Dunaja. Stúpnutie letných maxím teploty vody v Dunaji medzi Bratislavou a Medveďovom o 2º C zistené pred prehradením sa zachovalo. Ďalej sa zmenili hydrologické podmienky ako napríklad prietoky v ramennej sústave. Hlavné ramená sú od r. 1993 znovu trvalo prietočné, zvýšili sa rýchlosti prúdenia v hlavných ramenách, zmenil sa vzťah medzi unášacou silou, sedimentáciou a eróziou v ramenách, zväčšila sa výška hladiny vody a tým aj šírka ramien, predĺžila sa dĺžka a členitosť brehov, v depresiách vznikli nové stojaté vodné plochy, vzniklo meandrovanie zvyškového vodného toku v koryte Dunaja medzi opevnenými brehmi a pôvodné výhony v koryte Dunaja vytvárajú väčšiu variabilitu v prúdení vody. Hneď po znížení prietoku a výšky hladiny v pôvodnom hlavnom koryte v októbri 1992 sme na obnažených brehoch zaznamenali masový výskyt hynúcej bentickej makrofauny.
Na základe sledovania druhového spektra nálevníkov, pri hodnotení riečneho systému Dunaja ako celku, sa vplyv prehradenia prejavil výrazným nárastom druhového spektra po roku 1993 (Matis, Tirjaková, 1995b, Krno et al., 1999).
Zmeny sa výrazne prejavili aj na kvalitatívno-kvantitatívnom zastúpení jednotlivých druhov komárov (neustón). Vzhľadom na rýchlu reakciu druhov uvedenej čeľade na hydrologické zmeny, komáre sa javia ako veľmi vhodná indikačná skupina na monitorovanie prírodných zmien v skúmanom území. Spoločným znakom pre celú monitorovanú oblasť je výrazný pokles výskytu jarných druhov komárov najmä rodu Aëdes (A. vexans a A. sticticus) (Bulánková, Halgoš, 1995, Halgoš, Petrus, 1995), ako dôsledok absencie jarných záplav. Do spustenia VD bol v území dostatok podzemnej vody, čo sa prejavovalo i na zložení fauny komárov v priebehu roka. Pravidelné trojvrcholové krivky záplav, ktoré boli bežné v minulom období, sa zmenili na jednovrcholové krivky. Po sprevádzkovaní VD (Dobrohošť-Gabčíkovo) bol zaregistrovaný po r. 1993 výrazný pokles hladiny vody, kalamitné premnoženie komárov prestalo.
Spoločenstvá zooplanktónu po prehradení Dunaja
Eupotamál (hlavný tok)
V prvých dvoch rokoch po prehradení toku euplanktonické druhy v starom koryte v ročných priemeroch viac (pri perloočkách) alebo menej (pri veslonôžkach) ustúpili a naopak, tychoplanktonické (bentické a litorálne) formy nadobudli približne rovnaký kvantitatívny alebo vyšší podiel (Illyová, 1996, Vranovský, Illyová 1999). Z perloočiek to boli najmä druhy rodu Alona (A. quadrangularis, A. affinis a A. rectangula), Macrothrix hirsuticornis a Chydorus sphaericus, z veslonôžok Nitocra hibernica a Eucyclops serrulatus. Kvantitatívne uplatnenie nepravých planktontov bolo pritom (podobne ako pred prehradením Dunaja pri Čunove) zreteľnejšie v profile Dunajské kriviny ako v profile Gabčíkovo. Treba však pripomenúť, že pri jednotlivých odberoch kvantitatívny pomer dvoch porovnávaných ekologických skupín kolísal, a tiež, že zastúpenie pravých planktontov v hlavnom, resp. starom koryte Dunaja je závislé od ich prísunu najmä z bočných ramien typu parapotamál, prípadne plesiopotamál, situovaných nad daným profilom.
V potamoplanktóne derivovaného úseku Dunaja po prehradení pri Čunove došlo k určitému zníženiu priemerného podielu euplanktonických kôrovcov, čo je dôsledkom obmedzenia tvorby krustáceoplanktónu v ramenných sústavách a tým aj jeho obmedzeného vyplavovania do starého koryta (Vranovský, Illyová 1999)
Dunajské ramená a temporárne vody
Ramená typu parapotamál
Hydrologický režim ramien situovaných nad prístavom Gabčíkovo (patriacich k Vojčianskej, Šulianskej, Bodíckej a Bačianskej sústave) sa od sprevádzkovania VD značne líši od ramien medzi gabčíkovským prístavom a Sapom. Zatiaľ čo vyššie spomenuté ramená sa stali súčasťami umelo sprietočneného systému, napájaného z prívodného kanála pomocou odberového objektu pri Dobrohošti, ramená medzi Gabčíkovom a Sapom nie sú umelo dotované vodou. Na rozdiel od vyššie situovaných sú však relatívne intenzívne ovplyvňované vzdutím od sútoku starého koryta s odpadovým kanálom vodnej elektrárne. Tieto rozdiely sa od uvedenia odberového objektu Dobrohošť do prevádzky odrážajú aj na zložení krustáceoplanktónu. Tak v týchto ramenách vďaka ústupu pravých planktontov sa podstatne znížila abundancia krustáceoplanktónu a aj zooplanktónu, a zvýšil sa percentuálny podiel tychoplanktonických druhov: z perloočiek najmä viacerých druhov rodu Alona (hlavne A. guttata a A. rectangula), Chydorus sphaericus a Disparalona rostrata, z veslonôžok Nitocra hibernica. V roku 1995 naďalej pretrvávala nízka abundancia euplanktonických kôrovcov a väčšinou aj dominancia tychoplanktonických foriem (Vranovský, Illyová 1999).
V Istragovskom ramene, ktoré sa nachádza v oblasti nedotovanej vodou, zostal kvantitatívne relatívne bohatý krustaceoplanktón s pomerne silným zastúpením euplanktonických druhov, predovšetkým veslonôžok. Z euplanktonických perloočiek dominovali Bosmina longirostris alebo Diaphanosoma orghidani, z veslonôžok najčastejšie Cyclops vicinus, Acanthocyclops robustus alebo Thermocyclops oithonoides. Súvisí to s tým, že rameno nemá trvalo prietokový, ale väčšinou stagnantný režim, s výkyvmi v závislosti od vzdúvania hladiny v najspodnejšom úseku starého koryta. Zvýšenie podielu tychoplanktonických foriem v porovnaní s minulosťou je, podľa nášho názoru, spôsobené zrýchleným zabahňovaním, vyplytčovaním a zarastaním ramena.
V planktóne ramien typu parapotamál (bývalých slepých ramien) medzi obcou Dobrohošť a Gabčíkovom (rkm 1840-1820) sa po prehradení toku podstatne znížila priemerná početnosť a biomasa zooplanktónu, najmä planktonických kôrovcov. Percentuálny podiel euplanktonických kôrovcov výrazne klesol a dominanciu nadobudli nepravé, tychoplanktonické (litorálne a bentické) druhy. Uvedené zmeny sú dôsledkom skutočnosti, že ramenné sústavy - kde sa pred prehradením vďaka konektivite s hlavným tokom periodicky striedal stagnantný režim s prietokovým režimom, čo umožňovalo tvorbu krustaceoplanktónu v ramenách - boli premenené na trvalo prietočný systém, kde sú abiotické podmienky pre rozvoj zooplanktónu a najmä krustaceoplanktónu podstatne obmedzené. Takáto situácia v hlavných ramenách Vojčianskej až Šulianskej sústavy v podstate zotrvávala aj v roku 1995 a v nasledujúcich rokoch. Určité zvýšenie hodnôt početnosti a biomasy zooplanktónu, najmä krustaceoplanktónu v dolnej časti bývalej Bačianskej ramennej sústavy poukazuje na isté zlepšenie abiotických podmienok pre rozvoj zooplanktónu v najspodnejšom úseku prietočného systému (Vranovský a Illyová, 1999).
V bývalých ramenách typu parapotamál oblasti medzi Gabčíkovom a Sapom, napriek tomu, že ležia v derivovanej časti medzihrádzového územia, ostali aj po prehradení Dunaja zachované predchádzajúcemu stavu podobnejšie a pre rozvoj euplanktonických kôrovcov priaznivejšie podmienky. V priemere tu väčšinou naďalej dominujú euplanktonické druhy nad tychoplanktonickými.
Ramená typu plesiopotamál
V mediálnej zóne ramien typu plesiopotamál dominovali v taxocenózach kopepod euplanktonické druhy (najmä Eudiaptomus gracilis, Cyclops vicinus, Acanthocyclops robustus). V taxocenózach perloočiek v prvom období po prehradení prevládali litorálne, teda tychoplanktonické formy (najmä Chydorus sphaericus a Simocephalus vetulus) a tento trend zvyšovania percentuálneho podielu tychoplanktonických zástupcov pokračoval nielen pri perloočkách, ale aj pri veslonôžkach. V r. 1996-1997 tu dominoval z tychoplanktonických druhov Chydorus sphaericus a z euplanktonických Bosmina longirostris, z veslonôžok, v r. 1996, podobne ako v predchádzajúcom dominoval Thermocyclops oithonoides, v r. 1997 to bolo viac euplanktonických druhov. Medzi dominantné patril aj litorálny Eucyclops serrulatus ( Vranovský a Illyová (1999).
V bývalých ramenách typu plesiopotamál derivovaného úseku, pokiaľ neboli vysušené v dôsledku poklesu podzemnej vody, ostáva po prehradení druhovo pestré zoskupenie planktonických kôrovcov s pribúdajúcim podielom tychoplanktonických druhov, čo sa prejavuje aj zvýšeným indexom druhovej diverzity. Je to dôsledok prirodzeného procesu zarastania, starnutia a terestrifikácie týchto vodných útvarov, ktorý však urýchľuje znížená periodicita a intenzita umelých zátop v porovnaní so zátopami pred prehradením. V ramenách na juhovýchodnom okraji Bratislavy a tesne pod ňou (na obidvoch stranách zdrže), ktoré boli pred prehradením odsúdené na skorú terestrifikáciu, došlo vďaka vzostupu hladiny podzemných vôd k vytvoreniu podmienok pre revitalizáciu hydrocenóz vrátane zooplanktónu. Zvyšky ramien typu plesiopotamál, ktoré sa nachádzajú pod derivovaným úsekom, si zachovávajú hydrologický režim blízky režimu z obdobia pred uvedením VDG do prevádzky. Druhová bohatosť a diverzita ich krustaceoplanktónu má relatívne vysoké a postupne sa ešte zvyšujúce hodnoty. Súčasne však tieto plytké vodné útvary podliehajú prirodzenému procesu zarastania, vyplytčovania a zazemňovania (Vranovský, Illyová 1999).
Spoločenstvá zoobentosu a neustónu po prehradení Dunaja
Eupotamál (hlavný tok)
Vplyv prehradenia Dunaja sa prejavil výraznou zmenou spoločenstiev mikro- a meiozoobentosu (Matis, Tirjaková 1995 a, b). Zvýšil sa najmä podiel terestrických druhov v spoločenstve (zástupcovia rodu Colpoda). Tieto sa do toku dostali splavovaním po úpravách terénu zdrže. Výrazné zníženie prietoku po prehradení, výrazný pokles hladiny a naplnenie zdrže malo za následok formovanie spoločenstiev hnilobných procesov spojených s prudkým rozvojom baktérií, čo sa prejavilo nárastom abundancie a diverzity nálevníkov a bičíkovcov. Následkom týchto zmien došlo k zmenám v charaktere biotopu: od prudko tečúcich úsekov až ku spomaleniu toku a následnej izolácii niektorých lokalít, ktoré sa zmenili na stojaté vody s formovaním spoločenstiev typických stojatých vôd. V sprietočnených ramenách sa rozširuje druhové spektrum a abundancia najmä bičíkovcov a nálevníkov. Úsek nad prehradením (lokalita Kopáč) sa takmer nemení a počet druhov a nízka abundancia charakterizuje pôvodný tok. V tomto období sa tu vyskytli zriedkavé druhy, z nich niektoré boli zistené prvý raz na území Slovenska: Stentor multiformis, Balantidioides bivacuolata. V hlavnom toku z ostatných sledovaných skupín sa mierne zvýšila druhová diverzita i abundancia najmä Mastigophora, čo bolo spôsobené spomalením toku a zvýšenou sedimentáciou. Nápadné je zvyšovanie počtu terestrických druhov (Colpoda cucullus, C. inflata, Leptopharynx costatus, Colpoda steinii), čo svedčí o tom, že tok prináša mnoho sedimentov a dochádza k zanášaniu dna. Po prehradení nastáva ustaľovanie spoločenstiev s vysokou diverzitou ako aj početnosťou pričom sa spoločenstvá postupne čoraz viac podobajú spoločenstvám trvalo prietočných ramien. Od Dobrohošťa po Istragov postupne klesá druhové spektrum (Krno et al., 1999) a abundancia v dôsledku zmien dnového substrátu. Po napustení sa prejavila vyššia diverzita aj na lokalite Kľúčovec. Pred prehradením sa abundancia nálevníkov celoročne udržiavala na nízkych hodnotách, vplyv prehradenia sa prejavil až po roku 1993, keď dochádza k výraznému nárastu abundancie, ktorý sa udržiaval aj v nasledujúcich rokoch vzhľadom na stabilnejšie dno.
Medzi makrozoobentosom prevládajú zástupcovia permanentnej fauny a Chironomidae na úkor ostatných skupín temporálnej fauny. Zmena hydrologických podmienok spôsobila diferencovanie pôvodného koryta na dva rozdielne úseky. V hornom úseku kam, spadá profil pri Dunajských krivinách, znížením rýchlosti prúdu v koryte došlo k stabilizácii predtým pohyblivého dna. Zmenené abiotické faktory umožnili vytvorenie bohatých nárastov rias na pevnom štrkovitom dne. V dolnom úseku (Istragov), kde zasahuje spätné vzdutie, resp. Sporná sihoť, kde dochádza k pravidelnému dennému kolísaniu hladín, došlo k najväčšej zmene v štruktúre bentických biocenóz. Po úplnej zmene charakteru substrátu a hydrologických podmienok (výrazné zníženie prietoku a rýchlosti prúdu) došlo k deštrukcii pôvodného bentického spoločenstva. V prvom období bolo dno takmer bez zoobentosu a postupne dochádzalo k zvyšovaniu početnosti nového bentického spoločenstva (Krno et al., 1999). Po znížení prietoku a výšky hladiny v pôvodnom Dunaji v 1992 sme na obnažených brehoch zaznamenali masový výskyt hynúcej bentickej makrofauny.
Pri sledovaní permanentnej fauny sa po roku 1992 (Košel, 1995b, Krno et al., 1999) zaznamenal v hornom úseku (Dunajské kriviny - Gabčíkovo) nárast početnosti u skupiny Gastropoda (Košel, 1995a). K najväčšiemu zvýšeniu došlo u všetkých dovtedy dominantných druhov: Ancylus fluviatilis, Lymnaea ovata a Bithynia tentaculata. Ako nové druhy pribudli Nais christinae (Naididae), Limnodrilus hoffmeisteri, Potamothrix vejdovskyi (Tubificidae), Gammarus roeseli a Chaetogammarus tenellus (Amphipoda). Naopak poklesol podiel ploskule Dendrocoelum lacteum a kôrovca Dikerogammarus haemobaphes. Po roku 1995 v permanentnej faune dochádzalo k obohacovaniu druhového spektra, v Dunaji sa rozšíril ulitník Lithoglyphus. Podobne ako u temporálnej fauny dochádza k výraznému posunu v zastúpení potravných gíld od filtrátorov k zoškrabávačom (algofágom), čo odráža výrazný nárast perifytonu v rieke a prechod metabolizmu toku od heterotrofie ku autotrofii. Došlo tiež k zvýšeniu abundancie a biomasy pontokaspických druhov kôrovcov (Jaera istri, Corophium curvispinum a Dikerogammarus villosus), u máloštetinavcov dochádza k zvyšovaniu zástupcov čeľ. Tubificidae. Dominantnými sa preto stali druhy patriace k zoškrabávačom Dikerogammarus haemobaphes, Dikerogammarus villosus, Corophium curvispinum (Amphipoda), požierači vláknitých rias Jaera istri (Isopoda) a druhy z čeľ. Naididae žijúce na nárastoch rias. Usadený jemný sediment medzi štrkom sa stal vhodným biotopom pre druhy z čeľ. Tubificidae (Limnodrilus spp., Potamothrix sp.) osídľujúce bahnitý substrát. Zvýšené množstvo potravnej ponuky sa prejavilo u uvedených skupín v niekoľkonásobnom zvýšení ich abundancie a biomasy.
V druhovom spektre permanentnej bentickej fauny dolného úseku Dunaja (Istragove) došlo k zníženiu diverzity a vymiznutiu predtým dominatných rôznonožcov a objavenia sa nových zástupcov máloštetinavcov, ako druhy rodu Limnodrilus viazané na bahnitý substrát, pričom postupne sa zvyšovala ich abundancia. Ako nové sa tu objavujú stagnofilnejšie druhy Asellus aquaticus a Limnomysis benedeni (Crustacea). Po roku 1995 v profile Istragov došlo k vymiznutiu predtým dominantných rôznonožcov a objavenia sa nových zástupcov máloštetinavcov, ktoré sa tu predtým buď nevyskytovali alebo len v obmedzenej miere, ako druhy rodu Limnodrilus viazané na bahnitý substrát, pričom postupne u nich dochádzalo k zvyšovaniu abundancie. Po úplnej zmene charakteru substrátu a hydrologických podmienok (výrazné zníženie prietoku a rýchlosti prúdu) došlo k deštrukcii pôvodného bentického spoločenstva. V prvom období bolo preto dno takmer bez zoobentosu a až postupne dochádzalo k zvyšovaniu početnosti nového bentického spoločenstva s absolútnou prevahou máloštetinavcov s druhmi, ktoré nie sú pôvodnými pre tento typ biotopu Limnodrilus claparedeanus a Limnodrilus hoffmeisteri. V druhej polovici roka 1995 sme na Dunaji v úseku Dunajské kriviny - Gabčíkovo zaznamenali výraznejší pokles abundancie permanentnej fauny makrozoobentosu. Tieto zmeny súvisia so stavbou prehrádzky v koryte Dunaja pri Dunakiliti nad odberným profilom pri Dobrohošti, kedy došlo k silnému zákalu vody a prekrytiu dna jemnými splaveninami. V roku 1996 kolísanie výšky hladiny v koryte spôsobovalo zaplavovanie a obnažovanie ripálu toku v závislosti od zásobovania maďarskej ramennej sústavy vodou. Keďže však záplavové vlny mali krátke trvanie, bentická fauna v príbrežnej časti bola pomerne chudobná.
Podobne ako v regulovanom dolnom toku Rhôny (Fruget, 1991) prevládajú v Dunaji po roku 1992 v temporárnej faune (Tab. 5.8) (okrem pakomárov) Baetis fuscatus, Caenis luctuosa (Ephemeroptera), Psychomyia pusilla, Hydropsyche modesta a Ceraclea dissimilis (Trichoptera). Neskôr, napríklad v Rýne (Tittizer a kol., 1989), napriek zlepšeniu kvality vody k zásadnejším zmenám v štruktúre makrozoobentosu nedošlo. Súvisí to podobne ako na Dunaji s poklesom geomorfologickej diverzity rieky a jej prepojenia s ramennou sústavou, čo sa odráža na výraznom poklese heterogenity hydrosystémov (Fruget, 1992) a narušením funkčnej integrity prepojenia ramennej sústavy s riekou. Od roku 1993 došlo k výraznému kvalitatívnemu (Krno, 1995) a kvantitatívnemu ochudobneniu podeniek starého koryta, v ktorom sa pravidelne vyskytovali už len dva eurytopné druhy Baetis fuscatus a Caenis luctuosa (zriedkavejšie C. macrura). Oba tieto druhy nahradili druh Caenis pseudorivulorum a to v celom úseku od odklonenia Dunaja pri Dobrohošti až pod vyústenie vôd z Gabčíkovského kanála do Dunaja pri Sape. Podobne, fauna potočníkov bola zastúpená len dvoma rodmi - Hydropsyche a Psychomyia tak ako v regulovaných európskych veľtokoch Rhôna (Bournaud et al., 1990) a Rýn. Výrazne vzrástol podiel algofágov (Psychomyia) na úkor filtrátorov (Hydropsyche). Pričom druhy Hydropsyche contubernalis a H. bulgaroromanum boli nahradené druhmi H. pellucidula a H. modesta. (Krno et al., 1999). Neskôr sa tu objavil aj druh Rhyacophila dorsalis . Naposledy menované druhy sú typické pre menšie toky, akým sa Dunaj pod Dobrohošťom stal. Stabilizácia dna a lepšie trofické podmienky umožnili v 1994 a prvej polovici 1995 výrazný kvantitatívny nárast abundancie podeniek a najmä potočníkov.
Tab. 5.8 Zmeny v osídlení podeniek (Ephemeroptera), pošvatiek (Plecoptera) a potočníkov (Trichoptera) rieky
Dunaj v rokoch 1991-2001
Changes in population of Ephemeroptera, Plecoptera and Trichoptera in the Danube in 1991-2001
|
Taxón |
1991-92 |
1993-94 |
1995-96 |
1997-98 |
1999-00 |
2001- |
|
Ephemeroptera |
|
|
|
|
|
|
|
Leptophlebiidae |
|
|
|
|
|
|
|
Habrophlebia fusca (Curtis, 1834) |
+ |
|
|
|
|
|
|
Habrophlebia lauta (Eaton, 1884) |
+ |
|
|
|
|
|
|
Potamanthidae |
|
|
|
|
|
|
|
Potamanthus luteus ( Linneus, 1767) |
|
|
|
+ |
|
|
|
Ephemeridae |
|
|
|
|
|
|
|
Ephemera vulgata (Linneus, 1758) |
+ |
+ |
+ |
+ |
+ |
|
|
Caeniidae |
|
|
|
|
|
|
|
Caenis luctuosa (Burmeister, 1917) |
++ |
++ |
++ |
++ |
+ |
|
|
Caenis macrura (Stephens, 1835) |
+ |
++ |
+ |
+ |
|
|
|
Caenis pseudorivulorum (Keffermueller, 1960) |
+++ |
|
|
|
|
|
|
Ephemerelliidae |
|
|
|
|
|
|
|
Serratella ignita (Poda, 1761) |
|
+ |
|
|
|
|
|
Baetidae |
|
|
|
|
|
|
|
Baetis fuscatus ( Linnaeus, 1761) |
+++ |
+++ |
++ |
+ |
|
|
|
Baetis vardarensis (Ikomonov, 1962) |
+ |
|
|
|
|
|
|
Baetis vernus (Curtis, 1834) |
+ |
|
|
|
|
|
|
Centroptillum luteolum (Mueller, 1776) |
+ |
|
|
|
|
|
|
Cloeon dipterum ( Linnaeus, 1761) |
|
|
|
+ |
+ |
|
|
Procloeon bifidum (Bengtsson, 1912) |
|
|
|
+ |
|
|
|
Heptageniidae |
|
|
|
|
|
|
|
Heptagenia flava (Rostock,1877 ) |
++ |
|
|
|
|
|
|
Heptagenia sulphurea (Mueller, 1776) |
+++ |
++ |
+ |
++ |
|
|
|
Rhithrogena semicolorata (Curtis, 1834) |
|
|
|
+ |
|
|
|
Plecoptera |
|
|
|
|
|
|
|
Leuctridae |
|
|
|
|
|
|
|
Leuctra fusca (Linnaeus, 1758) |
|
+ |
|
+ |
|
|
|
Trichoptera |
|
|
|
|
|
|
|
Rhyacophilidae |
|
|
|
|
|
|
|
Rhyacophila dorsalis (Curtis,1834) |
|
+ |
|
|
+ |
|
|
Rhyacophila pascoei (McLachlan, 1879) |
|
+ |
|
|
|
|
|
Hydroptilidae |
|
|
|
|
|
|
|
Hydroptila sp. |
|
|
++ |
|
|
|
|
Hydropsychidae |
|
|
|
|
|
|
|
Hydropsyche bulgaromanorum (Malicky, 1977) |
+++ |
++ |
+ |
+ |
++ |
|
|
Hydropsyche contubernalis (McLachlan, 1865) |
+++ |
+++ |
++ |
++ |
|
|
|
Hydropsyche exocellata (Dufour, 1841) |
|
++ |
|
|
|
|
|
Hydropsyche incognita (Pitsch, 1993) |
+ |
++ |
++ |
+ |
+ |
++ |
|
Hydropsyche modesta (Navás, 1925) |
++ |
+++ |
++ |
+ |
++ |
|
|
Hydropsyche pellucidula (Curtis, 1834) |
+ |
++ |
+ |
++ |
+ |
|
|
Psychomyiidae |
|
|
|
|
|
|
|
Psychomyia pusilla (Fabricius, 1781) |
+++ |
+++ |
|
++ |
++ |
+ |
|
Brachycentridae |
|
|
|
|
|
|
|
Brachycentrus subnubilus (Curtis, 1834) |
+++ |
|
++ |
++ |
+ |
+++ |
|
Goera pilosa (Fabricius, 1775) |
|
|
|
|
+ |
++ |
|
Lepidostomatidae |
|
|
|
|
|
|
|
Lepidostoma hirtum (Fabricius, 1775) |
+ |
|
|
|
|
|
|
Leptoceridae |
|
|
|
|
|
|
|
Ceraclea dissimilis (Stephens, 1836) |
++ |
++ |
+ |
+ |
+ |
|
Vplyvom prehradenia došlo po roku 1992 na Dunaji k výraznému zvýšeniu počtu zaznamenaných druhov pakomárov (Krno et al., 1999). Kým v rokoch 1990-1992 sme tu registrovali 6 druhov, v roku 1993 až 1995 ich počet vzrástol na 18. Táto zmena súvisí so zmenou hydrologických podmienok, kedy v dôsledku zníženia prietoku a následného zníženia rýchlosti prúdu došlo k určitej stabilizácii dna, čo umožnilo osídľovanie viacerých nových druhov, ktorým predchádzajúce extrémne lotické podmienky nevyhovovali. Početnosť pôvodne dominantných druhov Chironomus sk. fluviatilis sa mierne zmenšila, kým početnosť niektorých pôvodne málo hojných druhov výrazne vzrástla (Microtendipes sk. chloris, Chironomus sk. reductus a Chironomus sp.). Medzi druhy, ktoré si zachovali svoju výskytovú i početnú stálosť patria Cryptochironomus sk. defectus a Dicrotendipes nervosus. Výrazný pokles rýchlosti prúdenia v dolnom úseku starého koryta Dunaja nad sútokom s kanálom od elektrárne spôsobil v oblasti Istragova vytvorenie vyše 0,5 m hrubej vrstvy piesčito-ílovitých usadenín, ktoré s výnimkou pakomárov (Chironomus sk. fluviatilis, Chironomus sk. reductus, Cryptochironomus sk. defectus, Endochironomus sp., Polypedilum sk. nubeculosum) a stagnikolného psamofilného druhu Ephemera vulgata (Ephemeroptera) nie sú osídlené vodným hmyzom.
Zatiaľ čo v Dunaji pred prehradením tvorila temporálna fauna v ročnom priemere okolo 30% z abundancie makrozoobentosu, v úseku ovplyvnenom VD klesá táto hodnota až pod 10% (rok 1998-2001), čo je istá analógia s jej prirodzeným zastúpením v dunajských ramenách. Na jeseň 2001 sa v osídlení potočníkov Dunaja veľmi pozitívne uplatnila umelá záplava. Opätovne sa objavil rod Hydropsyche, zastúpený druhom H. incognita, a viacero ďalších druhov potočníkov (Tab. 5.8).
Dunajské ramená a temporárne vody
Ramená typu parapotamál
V hornom úseku ramenného systému Dunaja (Bodícka brána), dotovanom vodou z prívodného kanála, so vznikom reofilných podmienok po napustení sústavy vodou sa druhová pestrosť mikro- a meiozoobentosu znížila. Spoločenstvá reagovali na zmenu charakteru ramena nestabilitou v oživení (Krno et al., 1999). V prietočných ramenách Dunaja bez vodnej dotácie (Istragov) je spoločenstvo mikro- a meiozoobentosu charakterizované typickými zástupcami sapropelu a bahnitého substrátu, ktorý je na celej lokalite dominantný (Krno et al., 1999). U permanentnej fauny osídľujúcej štrko-piesčitý litorál horného úseku prietočných ramien došlo na prelome rokov 1992-1993 k výraznému poklesu fauny máloštetinavcov v roku 1993, keď z bentickej fauny vymizli zástupcovia čeľade Tubificidae a Hypania invalida (Polychaeta). Z máloštetinavcov boli prítomné len druhy z čeľade Naididae osídľujúce perifyton so schopnosťou rýchlej rekolonizácie, prípadne amfibiotické druhy. Rekolonizácia ostatných skupín permanentnej fauny prebehla pomocou driftu. Rekolonizácia bola pomerne rýchla, čomu napovedá aj zvýšenie diverzity, keď sa táto zvýšila počas dvoch mesiacov takmer dvojnásobne. Podstatne nižšie zastúpenie mali vyššie kôrovce (Amphipoda, Mysidacea). V jarnom odbere, podobne ako v hlavnom toku Dunaja, dominovali z máloštetinavcov druhy z čeľade Naididae. V roku 1995 tu dominantné postavenie zaujímali druhy amfibiotické, pohyblivé (Amphipoda) a z máloštetinavcov druhy z čeľ. Naididae s krátkym životným cyklom. Neskôr dominantné postavenie na štrkovitom substráte z permanentnej fauny zaujímali Amphipoda. V roku 1997 sa druhová diverzita zvyšovala. Vysokú početnosť dosahovali v letnom a jesennom období Amphipoda. Vysokú početnosť dosahovala tiež Jaera istri (Isopoda).
Po roku 1992 stále prúdenie vody v umele napúšťaných horných
dunajských ramenách bodíckej sústavy umožňovalo výskyt reofilných druhov temporárnej
fauny. Litorálna zóna ramien je kolísaním hladiny vody často
obnažovaná, čo sa odzrkadľovalo na zmene vodnej fauny, ktorá ju
osídľuje (Krno et all.,
1999). V nových reofilných podmienkach, v ramenách dotovaných vodou je typicky
stagnikolný druh Caenis horaria
postupne nahradzovaný druhom C. luctuosa (Ephemeroptera).
Pravidelnejší výskyt druhov Baëtis
fuscatus, Caenis luctuosa (Ephemeroptera), Athripsodes cinereus, A. albifrons, Oecetis furva a rodu Anabolia
(Trichoptera) signalizujú trvalé napájanie ramien pôvodne typu plesiopotamál
povrchovými i podzemnými vodami. Naopak ustupujú typicky stagnikolné taxóny,
ako sú Caenis simile (Ephemeroptera),
rody Cyrnus, Oligotrichia a druh Mystacides azurea (Trichoptera). U vážok
v r. 1993 bol zaznamenaný výskyt pionierskych druhov Anax imperator a Libellula
quadrimaculata. V r. 1994-95 boli na lokalite zastúpené reofilné (Calopteryx splendens, Platycnemis pennipes)
i stagnofilné druhy: Enallagma
cyathigerum, Coenagrion puella. Zastúpenie spoločenstva pakomárovitých
v porovnaní s rokom 1992 sa zmenilo. Objavili sa nové druhy: Potthastia longimana, Paracladius conversus,
Endochironomus sk.signaticornis, Endochironomus
sp., Glyptotendipes sp., Polypedilum convictum a Micropsectra junci. Naopak,
nezaznamenali sme v minulých rokoch prítomné druhy Macropelopia nebulosa, Chironomus sk. fluviatilis, Ch. sk. salinarius,
Dicrotendipes sp., Paratendipes
intermedius, Stictochironomus sp.,
Paratanytarsus sk. lauterborni a Tanytarsus sp. V roku 1994 na lokalite pokračoval mohutný
nástup fytofilných druhov rodu Glyptotendipes
sp. a Polypedilum exectum, ktoré
v riasových nárastoch na štrkovitom substráte našli vhodné podmienky pre svoju
existenciu a sú najpočetnejšie zastúpené. V roku 1995 aj na základe
druhového spektra pakomárovitých možno konštatovať striedanie
hydrologických pomerov: na jar pri reofilných podmienkach (prepúšťanie
viac vody do ramennej sústavy) výskyt druhu Cricotopus
bicinctus, kým na jeseň (prepúšťanie menej vody) prítomnosť
druhov Microtendipes chloris, Glyptotendipes gripekoveni a Chironomus sk. thumi napovedala o tom, že hydrologické podmienky sa priblížili k
stojatým vodám, ktoré tieto druhy uprednostňujú. V prietočných
ramenách pribudlo po roku 1995 viacero prúdomilných druhov temporárnej fauny,
ide už o typické reofilné dunajské druhy Heptagenia
sulphurea, Serratella ignita a Psychomyia
pusilla (typické skôr pre ramená typu eupotamál). Vo všetkých ramenách
dotovaných vodou došlo k výraznému ústupu potočníka Ecnomus tenellus. Po roku 1995 v spoločenstve pakomárovitých
tu dominujú reofilné druhy. V jarnom období (Orthocladius thienemenni, Cricotopus bicinctus, Rheotanytarsus sp.,
Pottastia gaedii) a v druhej polovici
roka prevládali druhy Microtendipes
chloris, Glyptotendipes pallens a
G. gripekoveni uprednostňujúce
stojaté vody. (Krno
et al., 1999).
V ramenách bez vodnej dotácie (Istragov), kde nie je dosiaľ doriešené zavodňovanie ramien, došlo rovnako ako v hlavnom toku k výrazným zmenám v štruktúre vodnej fauny. V ramene v Istragove, ktoré bolo pôvodne prietočné a bez zárastov makrovegetácie sa začína objavovať sporadický výskyt submerznej makrovegetácie (Potamogeton pectinatus). Je to spôsobené tým, že do ramena podobne ako v sústave ramien v Spornej sihoti sa dostáva voda len cez výtokovú bránu (spätným vzdutím vôd vypúšťaných do hlavného toku VD Gabčíkovo) a nedochádza k pretekaniu vody cez hornú uzáveru ramena. V ramenách preto chýba prúd vody, ktorý pôvodne udržoval rameno bez vodnej makrovegetácie. Štrkovitý sediment je prekrytý bahnito-piesčito-ílovitým sedimentom s organickými zvyškami.
V ramene pri Istragove majú z permanentnej fauny dominantné postavenie máloštetinavce zastúpené druhmi z čeľ. Tubificidae (Limnodrilus hoffmeisteri, L. claparedeanus), ktoré tu dosahujú vysokú početnosť a biomasu. V roku 1997 sa po vysokých prietokoch v hlavnom toku v letnom období v odberoch objavili Amphipoda zastúpené druhmi Dikerogammarus bispinosus a Corophium curvispinum. V jesennom odbere sme ich prítomnosť už nezistili, čo potvrdzuje, že za terajších hydrologických podmienok v ramene nie sú pre nich vhodné životné podmienky.
Temporárna fauna (Tab. 5.9) v nich bola zastúpená hlavne larvami pakomárovitých. Najvýraznejšie zastúpenie tu majú druhy rodu Glyptotendipes, Chironomus sk. fluviatilis, Procladius sp., Cladotatytarsus sk. mancus. Čo sa týka početnosti jednotlivých druhov pakomárov nastal v roku 1995 jej radikálny pokles. Druhy rodu Glyptotendipes vypadli úplne a rodu Chironomus sa vyskytli len málo. Zastúpenie tu majú druhy Cricotopus sp., Procladius sp., Cladotatytarsus mancus na jar, Cryptochironomus defectus a Chironomus skup. semireductus. Aj u lariev vážok došlo k poklesu počtu druhov. Po roku 1996 nastali len mierne zmeny: 3 vymiznuté druhy (Cricotopus sp., Chironomus sk. thumi a Cryptochironomus sk. defectus) boli nahradené 2 novými druhmi (Endochironomus sk. nymphoides a Paratanytarsus skup. lauterborni). Nárast početnosti sme v porovnaní s rokom 1992 zaznamenali u druhov Chironomus skup. plumosus, Polypedilum sk. pedestre a najmä u dravého druhu Tanypus kraatzi. V nasledujúcich rokoch bolo spoločenstvo pakomárovitých stabilizované. Vyskytli sa aj akcesorické druhy Eukieferiella devonica, Brillia modesta a. Rheotanytarsus exiguus. Druhovo chudobne boli zastúpené vážky. Druhové spektrum vážok sa postupne zredukovalo z troch druhov vážok na jediný eurytopný druh Ischnura elegans elegans. Z hľadiska výskytu neustónu (Halgoš, 1995) sa uvedený biotop javí ako veľmi málo vhodný, počas uvedených rokov sa tu nachádzalo iba sporadicky niekoľko jedincov druhov Aëdes vexans a Aëdes sticticus. Zaujímavé je zistenie zo septembra v roku 1998 (Krno et al., 1999), kedy pri výraznom poklese úrovne hladiny vody v ramene spojeného so silným zárastom vodnej vegetácie bol zistený pomerne masový výskyt druhov Anopheles maculipennis s.l. a zriedkavo výskyt druhov Culex hortensis a Culex pipiens, čo svedčí o stabilnom charaktere.
Ramená typu plesiopotamál
Rameno pri Kráľovskej lúke (Foto 48-55), hoci leží v inundačnom území Dunaja nemá povrchové spojenie s okolitými ramenami. Zarastanie ramena makrovegetáciou pokračuje najmä po roku 1997, dochádza k masívnemu zarastaniu jeho voľnej hladiny a dna submerznou makrovegetáciou, čo spôsobuje jeho rýchle vyplytčovanie a zazemňovanie. Od začiatku až do septembra roka 1993 pretrvávala v ramene veľmi nízka hladina (0,3 m na vodočetnej late) a zavodňovanie ramennej sústavy sa prejavilo stúpnutím hladiny až po 7 mesiacoch v októbri. V období s nízkou hladinou teplota vody dosahovala až 33oC (9.6.1993).
Mikro- a meiofauna v tomto type ramien pokračuje v stabilizácii čo sa prejavuje aj vo zvyšovaní jej druhovej diverzity. Zo zriedkavo sa vyskytujúcich druhov tu boli zistené Histiobalantium natans, Tintinnopsis cylindrica, Ophrydium crassicaule, Frontonia ambigua, Holosticha grisea, Strombidium turbo, S. velox, Frontonia ambigua, Holosticha grisea a i. (Krno et al., 1999).
K typickým zástupcom permanentnej fauny patrili Asellus aquaticus (Isopoda) a Limnomysis benedeni (Mysidacea). Novými pre túto lokalitu sú druhy Psammoryctides albicola a Gammarus roeselii. V roku 1995 sa k typickým zástupcom zaradili Rhynchelmis limosella a Dero digitata a Nais spp. (Oligochaeta). Významne stúpla početnosť machoviek Plumatella fungosa a P. repens, ktoré pokrývali pevný substrát až na 50%. V roku 1994 sa po jednoročnej absencii objavili druhy, ktoré sa tu vyskytovali do roku 1992, napr. pijavica Erpobdella octoculata, lastúrniky Planorbis planorbis a Bithynia tentaculata. Objavili sa aj druhy predtým tu neznáme, ulitníky Physella acuta, Hippeutis complanatus. Došlo tiež k zvýšeniu kvantity u niektorých prežívajúcich ulitníkov, ako Lymnaea auricularia, L. stagnalis a Gyraulus albus. V roku 1995 v ramene sa zvýšila hladina vody oproti predchádzajúcim rokom. Na jar na voľnej hladine prevládali submerzné zárasty Batrachium sp., v letnom období Ceratophylum sp. a Myriophylum sp. Ako nový druh sa na lokalite zaznamenal ulitník Anisus vortex. V roku 1996 pribudli ploskuľa Dugesia lugubris (Turbellaria), pijavica Glossiphonia complanata (Hirudinea) a ulitník Armiger crista (Gastropoda). V roku 1997 sa objavili ďalšie nové druhy Chaetogaster langi a Pristina longiseta (nové pre faunu Slovenska) a pijavice Glossiphonia concolor, Theromyzon tessulatum a Erpobdella testacea.
Tab. 5.9 Zmeny v osídlení podeniek (Ephemeroptera), pošvatiek (Plecoptera) a potočníkov (Trichoptera) v dunajských ramenách v rokoch 1991-2001.
Changes in population of Ephemeroptera, Plecoptera and Trichoptera in the Danube river arms in 1991-2001
|
Taxón |
1991-92 |
1993-94 |
1995-96 |
1997-98 |
1999-00 |
2001- |
|
Ephemeroptera |
|
|
|
|
|
|
|
Ephemeridae |
|
|
|
|
|
|
|
Ephemera vulgata (Linneus, 1758) |
++ |
++ |
+++ |
++ |
++ |
|
|
Caeniidae |
|
|
|
|
|
|
|
Caenis horaria (Linnaeus, 1758) |
+++ |
+++ |
++ |
++ |
+ |
++ |
|
Caenis lactea (Burmeister, 1839) |
+ |
|
|
|
|
|
|
Caenis luctuosa (Burmeister, 1917) |
+++ |
+++ |
+++ |
+++ |
++ |
+++ |
|
Caenis macrura (Stephens, 1835) |
++ |
+ |
+ |
+ |
+ |
|
|
Caenis robusta (Eaton, 1884) |
|
++ |
++ |
++ |
++ |
++ |
|
Ephemerelliidae |
|
|
|
|
|
|
|
Serratella ignita (Poda, 1761) |
|
|
|
+ |
|
|
|
Baetidae |
|
|
|
|
|
|
|
Cloeon dipterum ( Linnaeus, 1761) |
+++ |
+++ |
+++ |
+++ |
+++ |
+++ |
|
Cloeon simile (Eaton, 1870) |
+ |
+ |
|
|
|
|
|
Heptageniidae |
|
|
|
|
|
|
|
Heptagenia flava (Rostock,1877) |
|
|
|
|
+ |
|
|
Heptagenia sulphurea (Mueller, 1776) |
|
|
|
+ |
|
|
|
Plecoptera |
|
|
|
|
|
|
|
Chloroperlidae |
|
|
|
|
|
|
|
Isoptena serricornis (Pictet, 1881) |
|
|
|
|
+ |
|
|
Trichoptera |
|
|
|
|
|
|
|
Hydroptilidae |
|
|
|
|
|
|
|
Agraylea sexmaculata (Curtis, 1834) |
|
+ |
|
|
|
|
|
Polycentropodidae |
|
|
|
|
|
|
|
Cyrnus trimaculatus (Curtis, 1834) |
|
+ |
|
|
|
|
|
Cyrnus flavidus (McLachlan, 1864) |
|
|
|
|
+ |
|
|
Holocentropus stagnalis (Albarda, 1874) |
|
|
+ |
|
|
|
|
Psychomyiidae |
|
|
|
|
|
|
|
Psychomyia pusilla (Fabricius, 1781) |
|
|
++ |
|
+ |
|
|
Ecnomidae |
|
|
|
|
|
|
|
Ecnomus tenellus (Rambur, 1842) |
+++ |
+++ |
+++ |
+ |
+ |
|
|
Phryganeidae |
|
|
|
|
|
|
|
Agrypnia varia (Fabricius, 1793) |
|
|
|
+ |
|
|
|
Phryganea bipunctata (Retzius, 1783) |
|
|
|
|
++ |
|
|
Limnephilidae |
|
|
|
|
|
|
|
Anabolia furcata (Brauer, 1857) |
+ |
|
++ |
++ |
|
++ |
|
Limnephilus flavicornis (Fabricius, 1787) |
|
++ |
+ |
+ |
|
|
|
Limnephilus lunatus (Curtis, 1834) |
|
++ |
+ |
|
|
|
|
Leptoceridae |
|
|
|
|
|
|
|
Athripsodes albifrons (Linnaeus, 1758) |
|
|
+ |
|
|
|
|
Athripsodes aterrimus (Stephens, 1836) |
|
|
|
+ |
+ |
|
|
Athripsodes cinereus (Curtis, 1834) |
++ |
++ |
++ |
+ |
+ |
+ |
|
Leptocerus tineiformis (Curtis, 1834) |
|
|
+ |
++ |
+++ |
|
|
Mystacides azurea (Linnaeus, 1761) |
|
++ |
+ |
|
|
|
|
Mystacides longicornis (Linnaeus, 1758) |
|
++ |
+++ |
++ |
|
|
|
Mystacides nigra (Linnaeus, 1758) |
|
+++ |
++ |
++ |
++ |
|
|
Oecetis furva (Rambur, 1842) |
|
+ |
|
|
|
|
|
Oecetis lacustris (Pictet, 1834) |
|
|
|
|
|
+ |
|
Oecetis ochracea (Curtis, 1825) |
|
|
++ |
|
|
|
U temporárnej fauny (Tab. 5.9) dochádza k zmene druhového zloženia podeniek. Caenis robusta, v týchto ramenách nahradzuje druhy C. horaria a C. luctuosa. Z vážok dominovali do roku 1995 v týchto lokalitách len stagnofilné druhy (Ischnura elegans, Coenagrion puella, C. pulchellum, Enallagma cyathigerum, druhy z rodu Erythromma). Zvyšujúcu sa eutrofizáciu a pretepľovanie vody potvrdil aj výskyt termofilných druhov Crocothemis erythraea, Sympetrum meridionale. Celkove bol pozorovaný u vážok zvyšujúci sa podiel stagnofilných druhov, ktoré prevládali nad eurytopnými. Chironomidae predstavovali charakteristické spoločenstvo pre stojaté ramená s vyvinutou makrovegtáciou, prípadne bohatými riasovými nárastami na štrkopiesčitom substráte (Procladius sp., Paracladius conversus, Endochironomus skup.nymphoides, Glyptotendipes sp. a Polypedilum convictum). Zníženie početnosti sme zaregistrovali u druhov Microtendipes chloris a Polypedilum pedestre. Novými druhmi tu boli po 1995 druhy Ablabesmyia monilis, Polypedilum sk. nubeculosum, Cladotanytatsus sk. mancus, Tanytarsus sk. obatifrons, Tanytarsus sk. macrosandalum, Paratanytarsus sk. lauterborni, Endochironomus sk. tendens a výrazný nárast druhov rodu Glyptotendipes. Po roku 1996 sa zvyšoval podiel stagnofilných druhov vážok, ktoré prevládali nad eurytopnými. Na spoločenstve pakomárovitých sa prejavovalo pokračovanie zarastania ramena makrovegetáciou a to výrazným zvýšením abundancie fytofilných a pelofilných druhov: Glyptotendipes gripekoveni, Einfeldia skup. pectoralis, Einfeldia skup. pagana, Tanytarsus skup. macrosandalum a Dicrotendipes nervosus.
Hydrobiologické pomery sa však po roku 1999 veľmi zhoršili. Boli obdobia, v ktorých sa epifauna vôbec nevyskytovala, najmä po zime (silné premnoženie makrofýt a ich následný rozklad spôsobil v ramenách toho typu takmer anoxické prostredie). Krátke júnové záplavy v júni 2001 spomalili nepriaznivý trend.
Zo zástupcov neustonu v týchto ramenách prevládali komáre (Bulánková, Halgoš, 1995): Anopheles maculipennis s.l., Culiseta annulata, Aëdes vexans, Aë. sticticus, Aë. cinereus, Aë. dorsalis, Aë. cantans, Aë. leucomelas, Ae. communis, Aë. flavescens, Culex pipiens, C. territans, C. modestus.
Jazierka v inundácii
Najväčší význam z hľadiska vývinu preimaginálnych štádií hlavne kalamitných druhov komárov v inundačnom území Dunaja, majú inundačné jazierka (Halgoš, 1995). Ide o prírodné depresie vzniknuté prirodzene alebo antropickou činnosťou, ktoré sú počas zvýšenia úrovne hladiny vody v hlavnom toku Dunaja veľmi silne podmáčané a zaplavované priesakovou podzemnou vodou. Z hľadiska monitoringu sme zvolili nasledujúce biotopy.
Na monitorovacej ploche Bodícka brána. v r. 1991 prevládali hlavne druhy rodu Aëdes a to Aë. vexans, Aë. sticticus, Aë. cinereus, v r. 1992 sme zistili výskyt druhov Culex pipiens, Culex territans a Anopheles maculipennis s.l. V ďalších rokoch začal biotop veľmi silne vysýchať. V roku 1995 bol vplyvom umelých záplav tento biotop výhodný pre vývin komárov. Zistili sa tu veľmi zaujímavé a dokonca veľmi vzácne druhy komárov, napr. Culex territans, Culex hortensis, Culiseta ochroptera. V roku 1996 sme zaregistrovali prienik priesakových vôd v neskorých jesenných mesiacoch vplyvom výrazného zvýšenia hladiny vody v hlavnom toku. Vplyvom umelých záplav v r. 1995-1997 (Krno et al., 1999), ako aj dobrými hydrologickými pomermi v hlavnom toku boli oba skúmané biotopy zaplavované priesakovou vodou, čo sa prejavilo i výskytom preimaginálnych štádií komárov, hlavne jarných druhov, napr. Aëdes cantans, Aë des communis a Aëdes leucomelas. Žiaľ, kladný vplyv umelých záplav bol eliminovaný výrubom lužného lesa a tým úplným zničením biotopov skúmaných lokalít.
V periodických vodách v oblasti Istragova bol v r. 1991-92 zaznamenaný výrazný rozvoj kalamitných druhov komárov. Od r. 1993 vplyvom VD a výrazným poklesom podzemných vôd začína silná degradácia tohto biotopu. Voda v ňom sa nachádzala iba počas atypicky zvýšenej hladiny vody v hlavnom toku, napr. v r. 1996-97 (bez prítomnosti preimaginálnych štádií komárov). V súčasnosti je biotop zarastený krovinatou vegetáciou a čiastočne zahrnutý odpadom z ťažby dreva.
Periodické vody
Na monitorovacej ploche Dunajské kriviny bol zistený v r. 1991 v mláke vzniknutej po daždi na lesnej ceste pomerne bohatý výskyt lariev druhu Anopheles maculipennis s.l. (Halgoš, 1995). Výskyt tohto druhu na takomto biotope je pomerne zriedkavý a v nasledujúcom období sa jeho výskyt v periodických mlákach nezaregistroval. V r. 1991 bol zvolený ako biotop periodická mláka závislá na dažďových zrážkach na monitorovacej ploche Istragov. Koncom júna sa tu zistil bohatý výskyt lariev a kukiel druhu Aëdes vexans. V budúcich rokoch hlavne vplyvom poklesu podzemných vôd, dažďové mláky veľmi rýchle vysýchali a neboli vhodné pre vývin preimaginálnych štádií komárov.
Ichtyocenóza v koryte Dunaja pod prehradením
Eupotamál (hlavný tok)
Po uvedení VD Gabčíkovo do prevádzky došlo v starom koryte k značnému zníženiu prietoku vody, spomaleniu rýchlosti prúdu, posunutiu brehovej línie do stredu riečiska a vyplytčeniu, resp. na lokalite Istragov i k zabahneniu litorálu, ktorý je bez prirodzených úkrytov pre ryby. Zmeny sa prejavili v znížení relatívnej početnosti a druhovej diverzity ichtyocenózy. Zistené druhové spektrum sa znížilo z 19 druhov zistených v r. 1991-92 na 7 druhov v r. 1993. Nedostatok úkrytov a zabahnenie litorálu spôsobili, že túto lokalitu nevyhľadávajú druhy reofilné, ale eurytopné. Nedostatok úkrytov a monotónnosť litorálu zapríčinili, že sa zredukovala druhová diverzita. Z druhov, ktoré sa po roku 1993 nezistili patrilo 9 do skupiny reofilných (L. lota, G. baloni, B. barbus, C. gobio, L. cephalus, L. idus, G. kessleri, G. albipinnatus L. leuciscus, L. idus a B. barbatulus) a 8 do skupiny eurytopných druhov (A. alburnus, A. bjoerkna, C. auratus, C. carpio, E. lucius, S.lucioperca a P. fluviatilis, P. marmoratus [Černý (1999)].
Z hľadiska ohrozenia vymizli kriticky ohrozené druhy (divoká forma Cyprinus carpio a Gobio kessleri) ohrozený druh (Gymnocephalus baloni) a druhy vyžadujúce pozornosť (I - L. idus a L. lota). Znepokojujúci je však i ústup populácií druhov Chondrostoma nasus a Stizostedion lucioperca.
Dunajské ramená
Parapotamál:
V ramene pri Bodíckej bráne boli zaznamenané kvantitatívne zmeny v zložení ichtyocenózy, menovite v zmenšení podielu hospodársky cenných druhov a najmä predátorov. Vysvetľuje sa to okrem hydrologických zmien súvisiacich s výstavbou VD Gabčíkovo hlavne ilegálnym lovom rýb. Ilegálny odlov, ktorý zdecimoval ichtyocenózu bodíckej sústavy sa masovo realizoval najmä na jeseň r. 1992, v čase veľmi nízkych vodných stavov a tiež v období presmerovania hlavného toku Dunaja do derivačného kanála. Výraznejšie zmeny charakteru monitorovacej plochy Bodícka brána zmiernilo dotovanie tejto sústavy vodou z derivačného kanála. Pokles abundancie a druhovej diverzity rýb (z 19 druhov v r. 1990 na 8 druhov v r. 1994) na tejto lokalite nastal najmä v čase prehradenia hlavného toku Dunaja, kedy, pri zníženom vodnom stave, podstatná časť ichtyocenózy emigrovala a časť bola v terénnych depresiách pytliaci vylovili. Po vybudovaní prepážok v ramennej sústave sa síce zamedzila spätná imigrácia ichtyofauny z hlavného toku, avšak po dotovaní ramien vodou z derivačného kanála badať v tomto ramennom systéme určitú stabilizáciu zloženia ichtyofauny (Černý 1999).
Plesiopotamál
Rameno pri Kráľovskej lúke sa v r. 1981-90 vyznačovalo vysokou abundanciou ichtyocenózy. V r. 1991 sa však zistilo, že kolísanie vodného stavu v ramene sa značne oneskoruje za kolísaním hladiny v hlavnom toku. Táto skutočnosť posilňovala proces zazemňovania a starnutia ramena. Tento proces sa okrem iného značne urýchľoval tým, že v starom koryte Dunaja po spustení VD do prevádzky bol podstatne nižší prietok. Rameno nebolo dotované z hlavného toku tak ako v minulosti, naopak staré koryto Dunaja čiastočne spôsobovalo drenážny efekt. Hladina vody v ramene sa znížila natoľko, že vyschli plytké časti ramena porastené Nuphar luteum a Trapa natans. V prípade Kráľovskej lúky je tento proces stárnutia ramena rýchlejší i z dôvodu, že rameno bolo prevážnu časť roka typu plesio-paleopotamon a leží na okraji najširšej oblasti inundácie na našom území. Vyššie uvedené faktory spolu s intenzívnym lovom, najmä dravcov a hospodársky cenných druhov rýb, znižovali druhovú diverzitu ichtyocenózy v tomto ramene. V obsádke vzrástla početnosť ekologicky plastických, eurytopných druhov (Rutilus rutilus), introdukovaných (Lepomis gibbosus) a expanzívnych druhov (Carassius gibelio) a znížila sa relatívna početnosť ohrozených druhov, ktoré v minulosti patrili síce medzi recedentné druhy, avšak pravdepodobne v priebehu r. 1992 z ramena vymizli. U sazana (Cyprinus carpio) a zubáča volžského (Sander volgense) je tento stav zapríčinený pravdepodobne rybárskym tlakom, u ohrozených druhov (Gymnocephalus schraetser) a vzácnych druhov (Abramis sapa) zamedzením komunikácie ramena s ostatnými vodami inundačného územia Černý (1999).