2. Charakteristika územia
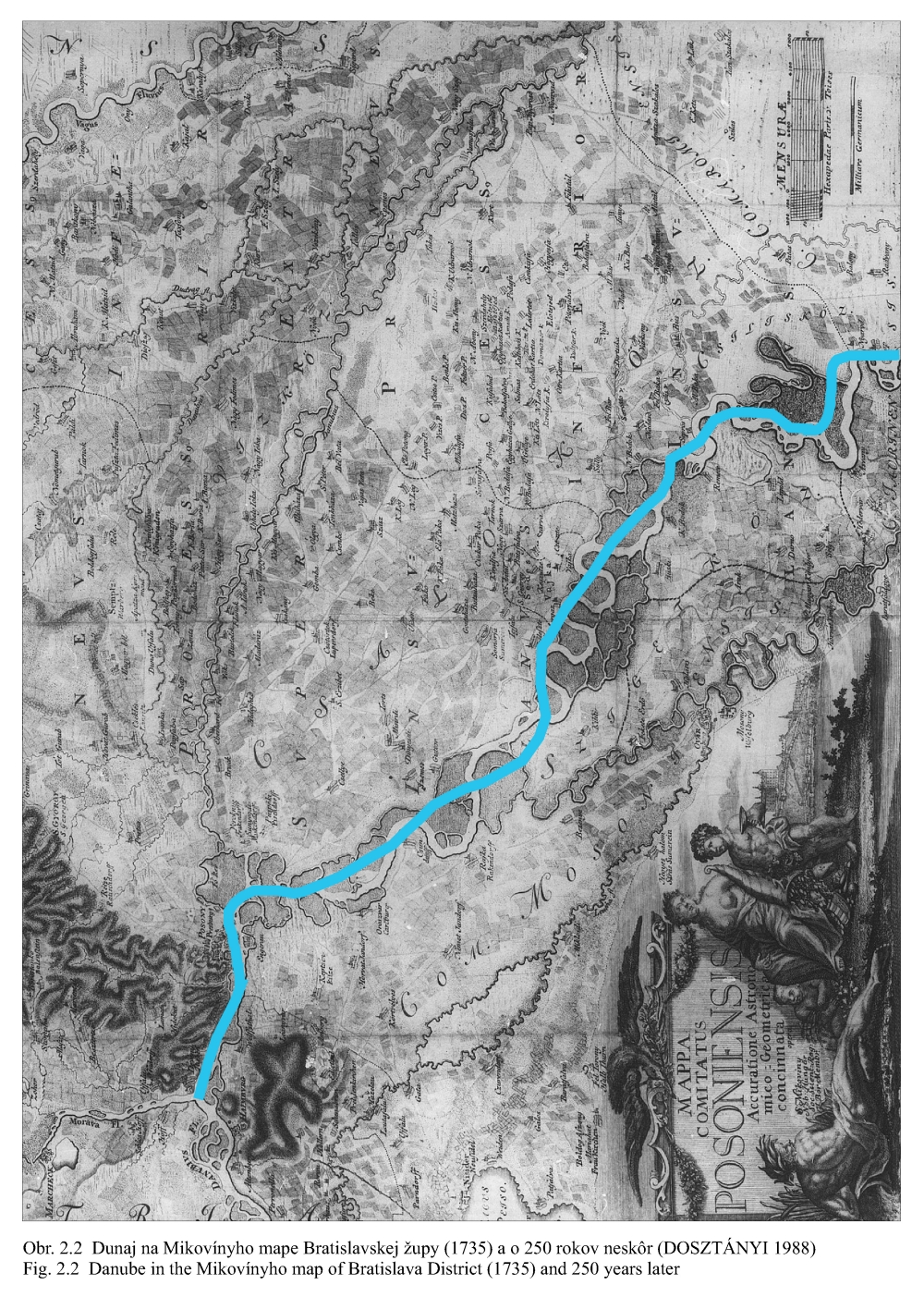
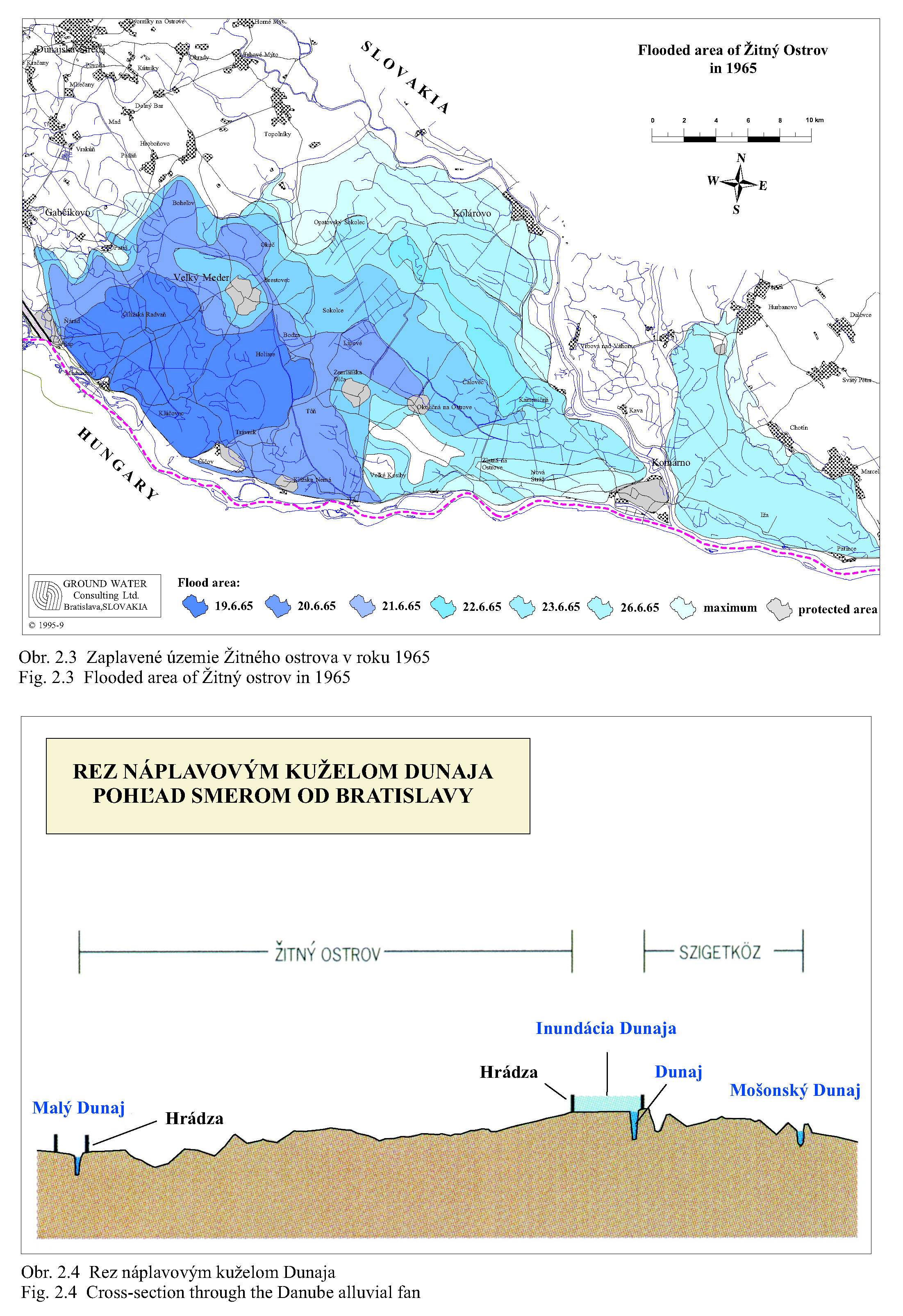
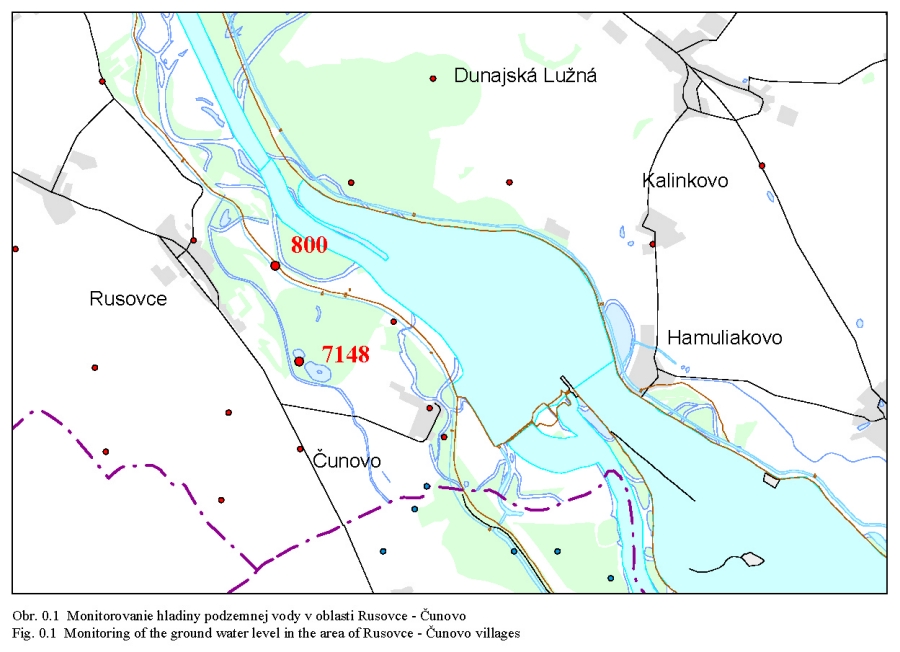
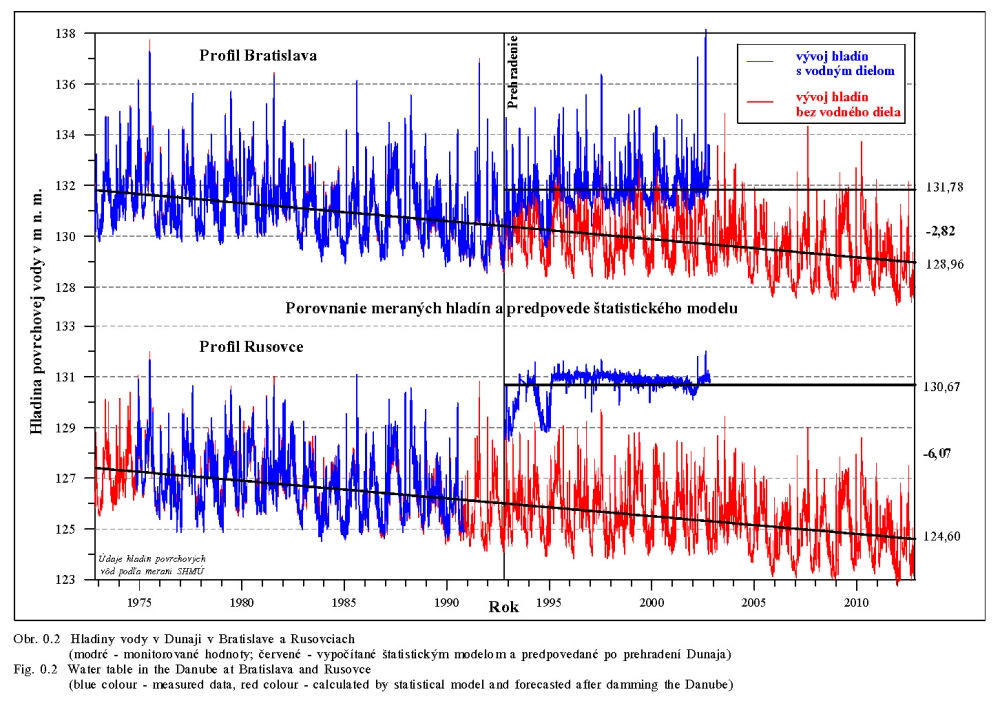
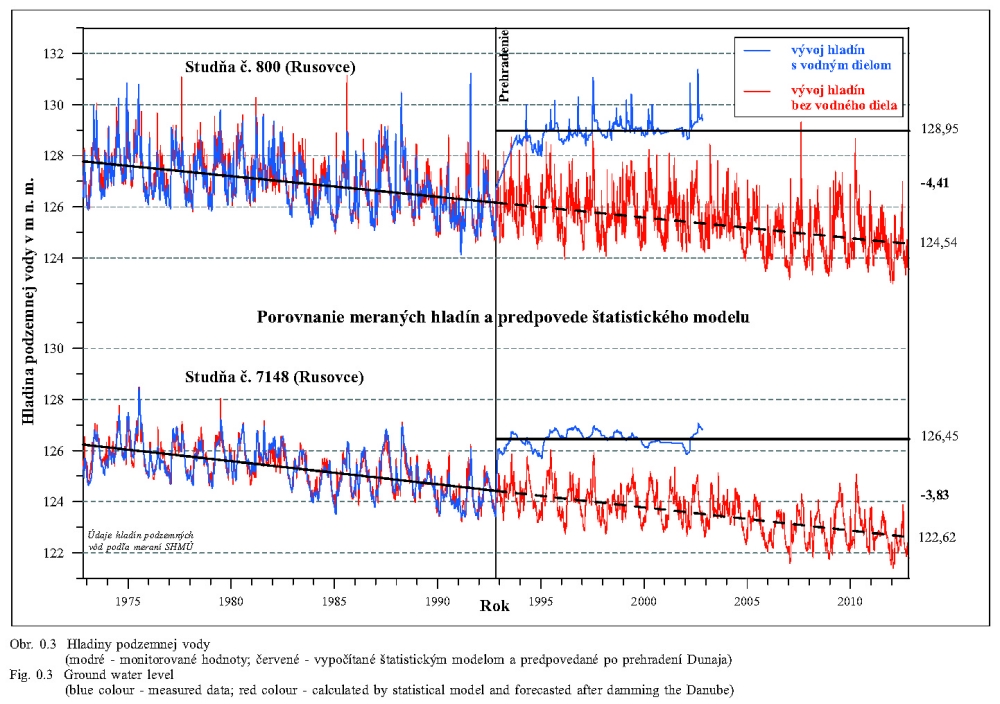
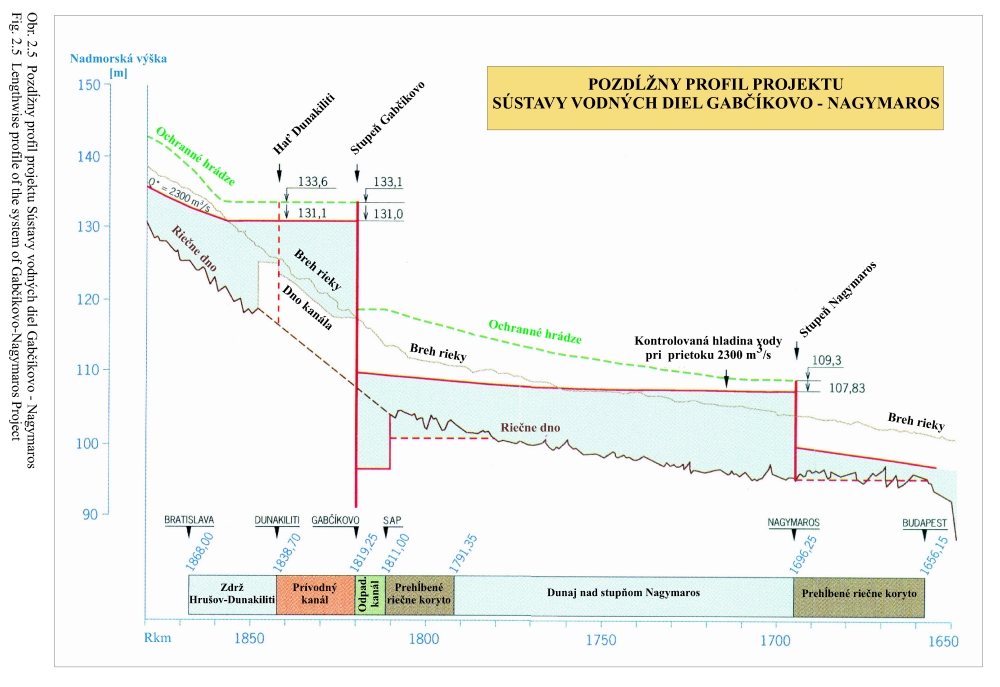
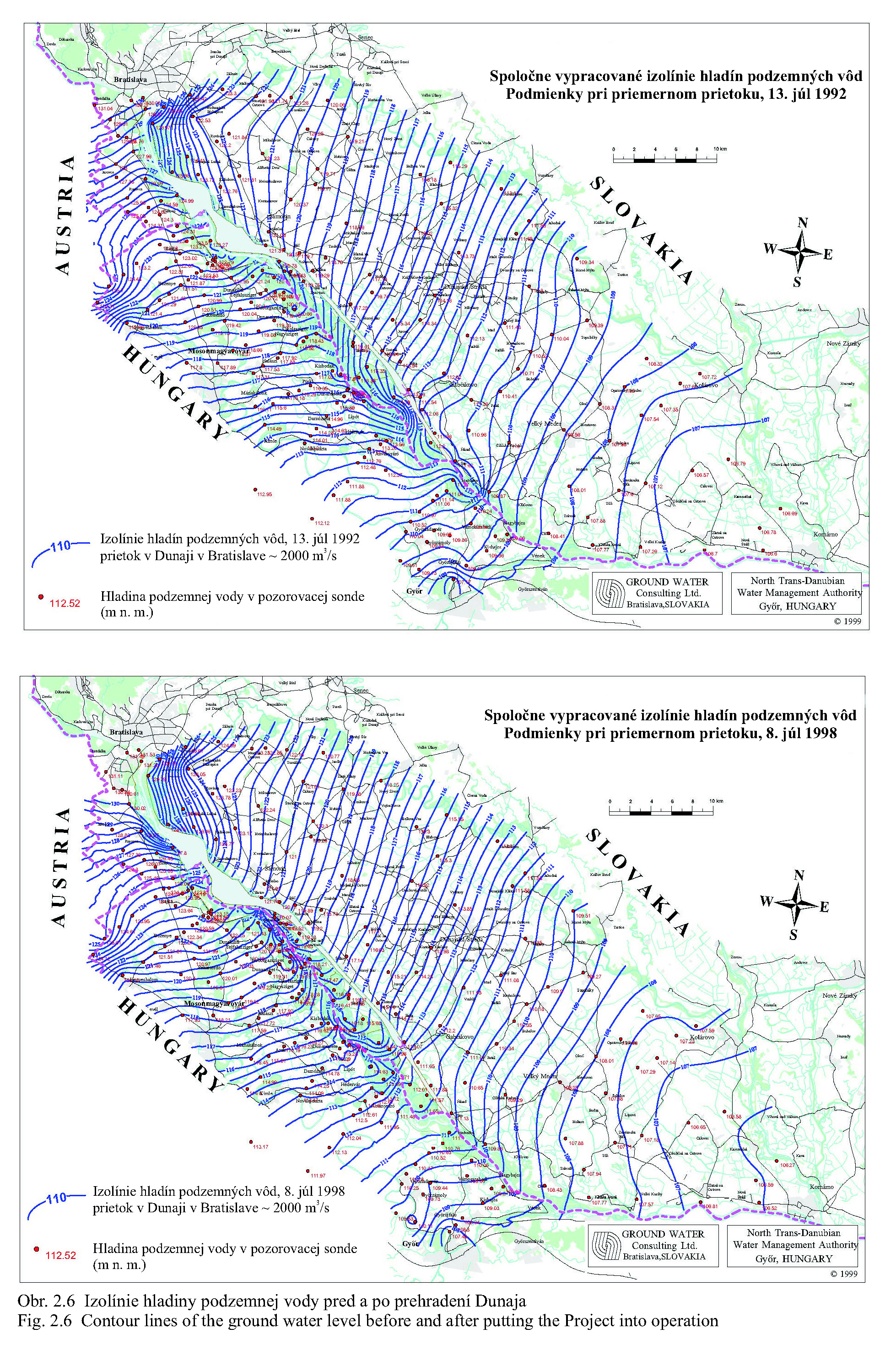
Posudzovaným územím optimalizácie vodného režimu je staré koryto Dunaja a jeho ľavostranné inundačné územie s ramennou sústavou od obce Dobrohošť (rkm 1842) po Sap (rkm 1811) (Obr. 2.1). Inundačné územie leží od Dunaja po pôvodné ochranné protipovodňové hrádze. Historické zmeny v dunajskom prírodnom prostredí sú výsledkom geologického vývoja a častých zmien klimatických pomerov vo štvrtohorách. Treba sem zahrnúť aj intenzívny pohyb štrkov a pieskov v Dunaji, prehlbovanie a dvíhanie riečneho koryta, tvorenie a putovanie riečnych meandrov (kľukatenie rieky), ukladanie nánosov, eróziu a zmeny brehovej čiary a časté záplavy územia. Prvá komplexná etapa budovania ochranných protipovodňových a regulačných opatrení na Dunaji sa realizovala v rokoch 1759-1914. Hlavná plavebná kineta, ktorá sa v zásade odvtedy používa, sa vytvorila reguláciou toku, ktorá začala v r. 1831 a bola dokončená v posledných rokoch 19. storočia (Obr.2.2). Dnešné vedenie protipovodňových hrádzí a neprirodzené, kvôli plavbe napriamené koryto, boli realizované po povodni v roku 1853. Tak vzniklo dnešné inundačné územie, o ktorom v tejto štúdii hovoríme. Povodeň v roku 1954 po pretrhnutí pravostrannej hrádze zdevastovala veľkú časť ostrova Szigetköz v Maďarsku (Dub 1954). O rozsahu katastrofy svedčí fakt, že polovica Szigetköz bola zaplavená a voda v Bácsi (okres Győr) stúpla až do výšky okien prvého poschodia domov. Povodeň v roku 1965 zaplavila dolnú časť Žitného ostrova na Slovensku. Plocha zaplaveného územia v roku 1965 bola 71700 ha poľnohospodárskej pôdy, 114000 ha pôdy bolo zamokrených, bolo zničených 3910 domov a evakuovaných 53693 obyvateľov (Hronec, 1969), (Obr. 2.3). Protipovodňová ochrana územia pod Bratislavou bola zahrnutá do projektu Gabčíkovo-Nagymaros. Dunaj veľmi prispel k rozvoju podunajských štátov. Je to rieka, ktorá bola intenzívne využívaná lodnou dopravou, pre zásobovanie vodou, na lovenie rýb, vodou pre poľnohospodárstvo, inundáciou pre pestovanie lesov, a od pradávna aj ako strategická vojenská hranica. Dunaj je od prvej polovice minulého storočia využívaný aj na výrobu elektrickej energie a na získavanie iných úžitkov. Niet pochybností o tom, že každé takéto využitie Dunaja, ale aj každá prírodná zmena, vplýva na prírodné prostredie. Rovnako niet pochybností o tom, že ak sa zasiahne do toku, ako sa to stalo v projekte výstavby Sústavy vodných diel Gabčíkovo – Nagymaros, vplýva to na prírodné prostredie, z niektorého hľadiska pozitívne a z iného negatívne. Toto platí pre všetky projekty a ľudské činnosti. Moderná technológia, ktorá umožňuje realizovať komplexné vodohospodárske projekty, obsahuje aj metodiku monitorovania, databázového spracovania, interpretácie a hodnotenia vplyvu na prírodné prostredie, s cieľom vyhnúť sa, kompenzovať, zmenšiť alebo napraviť takéto nepriaznivé vplyvy. Slovenskí ekológovia spolu s ochrancami prírody ako aj neskôr nezávislí experti EÚ (Európskej únie) (CEC 1992) vo svojej správe o zistených faktoch konštatovali, že „redukovanie prietoku v Dunaji má nepriaznivý vplyv na prírodné prostredie, pokiaľ nebudú realizované správne opatrenia“ (Foto 3, Foto 4). Rozsudok Medzinárodného Súdneho Dvora (International Court of Justice, 1997) konštatuje, že Projekt Sústavy vodných diel Gabčíkovo - Nagymaros „nebol len spoločným investičným projektom na výrobu energie, ale slúžil aj iným cieľom, ako zlepšenie plavby na Dunaji, ochrana pred povodňami, prevádzanie ľadov a ochrana prírodného prostredia”. Derivačný kanál bol postavený mimo inundačného územia (na rozdiel od všetkých ostatných priehrad na Dunaji) (Obr. 2.1). Nezávislí experti Pracovnej skupiny Komisie Európskeho spoločenstva vo svojej správe z 23. novembra 1992 (CEC 1992) k tomu konštatovali: "V minulosti vykonané zásahy pre plavbu obmedzili možnosti pre rozvoj Dunaja a jeho inundačného územia. Za predpokladu, že hlavné riečne koryto sa nebude viac používať na plavebné účely, vznikla unikátna situácia. S podporou technických opatrení sa rieka a záplavové územie môže vyvíjať prirodzenejšie ". Okrem toho na oboch stranách Dunaja boli vybudované objekty na trvalé zásobovanie Malého Dunaja, Mošonského Dunaja a ramenných sústav vodou. Predpokladalo sa, že hladina vody v starom koryte Dunaja sa technickými opatreniami udrží na úrovni podobnej akú mal Dunaj pred prehradením pri prietokoch okolo 1400 – 1500 m3/s. Starým korytom Dunaja sa dnes nazýva úsek Dunaja medzi Sapom, rkm 1811,0 až Čunovom, rkm 1851,75, o dĺžke 41,75 km, v ktorom po prehradení koryta pri Čunove chýba prevažná časť prietokov odvedená derivačným kanálom na vodnú elektráreň Gabčíkovo (Obr. 2.1). V súčasnosti starým korytom Dunaja preteká prietok v zmysle Dohody z roku 1995 od 250 do 600 m3/s vody, a počas povodní podstatne viac. V správe Trojstrannej komisie na zistenie faktov Európskeho spoločenstva z 31. októbra 1992 sa objavuje formulácia: "neuvedenie sústavy do prevádzky by viedlo ku značným finančným stratám a viedlo by ku vzniku vážnych environmentálnych problémov" (FFM, 1992). Tieto environmentálne problémy vychádzali zo zistenia, že v dôsledku predchádzajúcich úprav na Dunaji, hlavne kvôli zabezpečeniu plavby pri malých prietokoch a pre zabezpečenie protipovodňových opatrení, koryto Dunaja postupne poklesávalo a poklesávala aj hladina podzemnej vody (Obr. 0.1, Obr. 0.2 a Obr. 0.3). Tieto závery, ako aj ďalší pokles hladín podzemných vôd a vysýchanie ramennej sústavy, so všetkými negatívnymi dôsledkami na prírodné prostredie, bez vybudovania Vodného diela Gabčíkovo, sa potvrdili aj v štúdii Prognóza stavu hladín povrchových a podzemných vôd v podmienkach bez vybudovania vodného diela Gabčíkovo (Mucha, Banský, et al., 2001). Podľa expertov Komisie Európskeho spoločenstva (CEC 1992) prietok vo všetkých riečnych ramenách existoval pred prehradením v priemere iba počas 17-tich dní v roku. Gabčíkovská časť Sústavy vodných diel Gabčíkovo-Nagymaros leží v centrálnej časti medzihorskej depresie, v dunajskej kotline, nazývanej Podunajská nížina (Danubian Lowland). Dunajská kotlina je vyplnená usadeninami mladších treťohôr (morskými a jazernými pieskami, jemným pieskom, ílom, pieskovcom a bridlicami) a štvrtohôr (od mindelu už dunajským riečnym pieskom a štrkom sedimentujúcim v aluviálnych podmienkach Dunaja) usadzujúcich sa v riečnych alebo jazerných podmienkach. Celková hĺbka štvrtohorných a treťohorných usadenín je až 8000 m, pričom vrchné dunajské riečne sedimenty (od mindelu) vytvárajú hlavnú zvodnenú vrstvu zloženú z vysoko priepustných štrkov a pieskov. Hrúbka dunajských riečnych sedimentov sa pohybuje od niekoľkých metrov pri Bratislave, do viac ako 450 metrov pri Gabčíkove. Ďalej po prúde pod obcou Sap, v smere na Komárno, opäť klesá na hrúbku niekoľkých metrov. Pod touto vrstvou sa nachádza súvrstvie málo až takmer nepriepustných starších kvartérnych (pred mindelom) a hlavne treťohorných sedimentov. Dôležitým faktorom pri transporte dunajských sedimentov je existencia žulového prahu medzi Alpami a Karpatami v oblasti Bratislavy, s výstupmi žuly do riečneho dna. Podobné kamenné, predovšetkým andezitové, prahy sa nachádzajú aj od miest Štúrovo-Ostrihom po Visegrád-Nagymaros, približne 160 km pod Bratislavou. Obidva skalné prahy predstavujú pre vodný tok prírodné geologické bariéry, stupne alebo prahy. Tesne pod Bratislavou, pod žulovým riečnym prahom, sa od Dunaja oddeľujú toky dvoch jeho ramien - Malého Dunaja na slovenskej strane a Mošonského Dunaja na maďarskej strane. Tieto dve ramená ohraničujú spolu s Dunajom dva podobné ostrovy - Žitný ostrov na Slovensku a Szigetköz v Maďarsku. V tomto priestore, do ktorého je situovaná gabčíkovská časť projektu Sústavy vodných diel Gabčíkovo-Nagymaros, medzi Bratislavou a Medveďovom, Dunaj v minulosti vytvoril "vnútrozemskú deltu" (v geologickej literatúre opisovanú ako náplavový kužeľ). V tomto priestore Dunaj kedysi meandroval a rozvetvoval sa a usadzoval náplavový kužeľ, približne od Malého Dunaja až za Mošonský Dunaj. Táto vnútrozemská delta má svoju svojskú morfológiu: riečne meandre, akumuláciu hrubozrnných štrkov a pieskov a ich eróziu, zmeny sklonu riečneho dna a podobne. Tento veľký náplavový kužeľ pozostáva z extrémne priepustných a rozsiahlych zvodnených vrstiev, schopných zachytávať a transportovať veľké množstvá podzemnej vody. Dunaj tečie na povrchu tohoto kužeľa (Obr. 2.4). Voda z Dunaja preniká do náplavového kužeľa a prúdi ako podzemná voda pozdĺž Dunaja a smerom k Malému a Mošonskému Dunaju. V dolnom úseku, tam kde sklon rieky zrazu klesne na štvrtinu oproti sklonu pri Bratislave (Obr. 2.5), podzemná voda prúdi späť do Dunaja cez jeho dno, ramená, prítoky a odvodňovacie kanále (Obr. 2.6). Všetko toto je výsledkom zníženej priepustnosti a hrúbky zvodnenej vrstvy poniže Gabčíkova, čo je zase spôsobené tým, že andezitový prah pri Nagymarosi spomalil prúdenie a zmenil nad sebou sedimentačné podmienky Dunaja a k ním sa pripojila aj sedimentácia hlavne rieky Váh (Kolárovská formácia). Na tomto úseku Dunaja, od Bratislavy po Sap, boli jeho brehy opevnené a po oboch jeho stranách boli v minulosti vybudované protipovodňové hrádze medzi ktorými tečie napriamený Dunaj a existuje aj časť dunajských ramien (Obr. 2.1). Aj v súčasnosti sa počas vysokých prietokov dostane voda z Dunaja až k týmto hrádzam. Z prírodného a ekologického hľadiska sa územie považuje za veľmi cenné, unikátne a hodné zachovania jeho pôvodných funkcií. Okrem toho má toto inundačné územie aj nenahraditeľné funkcie ako je prevádzanie povodňových prietokov, prirodzenú funkciu poldra, ktorou sa znižujú maximálne prietoky počas povodní, čo sa významne prejavuje aj v nižších maximálnych prietokoch Dunaja v smere toku. Územie má vhodné podmienky na rast prirodzených lužných lesov i komerčnú produkciu dreva. Je esteticky i turisticky atraktívne a v prípade, ak v riečnych ramenách tečie voda, zabezpečuje aj funkciu „samočistiacich procesov“ čistenia riečnej vody. Po uvedení nápustného objektu do ramennej sústavy inundačného územia pri Dobrohošti do činnosti sa hydrologický režim priblížil stavu ku koncu 60-tych rokov. Bolo to konštatované po porovnaní dynamiky hladiny podzemných vôd z roku 1953 (Jurko 1958) s rokmi 1992-2000 (Mucha et al., 2001). Neplatí to o drénovanom páse pozdĺž starého koryta a o celom komplexe lesov pod gabčíkovským prístavom. V súhrne preto možno konštatovať, že napriek ozdravným hydropedologickým opatreniam je vodný režim sčasti nepôvodný. Ide približne o 30 % územia lužných lesov, kde je tento stav možné zlepšiť dnovými prehrádzkami, variabilitou prietokov a osobitným režimom zavodňovania širšieho okolia Istragova. Pôvodné vŕbovo-topoľové lužné lesy, ktoré boli dominantnou časťou vegetácie tohto územia, majú približne z 80 % ich rozlohy zmenenú drevinovú skladbu v prospech rozmanitých šľachtencov topoľov, menej aj vŕb. Len 15 – 20 % rozlohy týchto lesov má prírodnú drevinovú skladbu a to hlavne v menej dostupných porastoch (ostrovčeky, depresie). Bylinný podrast väčšiny dospelých (25-30 ročných) topoľových monokultúr nevykazuje veľké odchýlky od pôvodného floristického zloženia. Potvrdilo to porovnávanie fytocenóz pôvodného lesa a monokultúr urobené v roku 2001 v polesí Gabčíkovo (Krajňáková 2001). Bylinný porast je poznamenaný inváziou nepôvodných druhov z rodov Aster, Solidago a Impatiens, pričom neprítomnosť, resp. nedostatočnosť záplav túto inváziu podporuje. I keď o nelesnej vegetácii existuje len veľmi málo starých podkladov, môžeme ju, pokiaľ ide o brehové jednoročné spoločenstvá riečneho litorálu považovať za pôvodnú (prírodnú), kým kosené lúky a pasienky sú pochopiteľne antropogénne. Pre detailné posúdenie bola vypracovaná podrobná fytocenologická mapa (1:25000 a 1:10.000) (Šomšák et al., 2001, 2002, 2003), ktorá je podkladom pre všetky odbory bioty, ale aj pre posúdenie vplyvu existujúceho, či optimálneho vodného režimu. Táto mapa (Šomšák et al., 2003) je tiež pracovným podkladom pre rokovania o implementácii Rozsudku Medzinárodného Súdneho Dvora (International Court of Justice, 1997). 2.4 PRÍRODNOSŤ, FUNKČNOSŤ A ODVODENOSŤ EKOSYSTÉMU Pri posudzovaní kvality súčasného stavu akéhokoľvek ekosystému a vývoja, akým k nemu dospel, musíme rozlišovať prírodnosť, prirodzenosť a antropicky podmienenú, alebo priam antropogénne vzniknutú umelosť. Pri posudzovaní ekosystému rieka-zaplavované územie treba okrem toho vedieť, že ide o azonálny ekologický systém, v ktorom je rozhodujúcim faktorom hydrologický režim. Aj keď sa názory odborníkov líšia pri posudzovaní toho čo možno považovať za prírodné, teda pôvodné a človekom neovplyvnené a do akej miery je pri antropických zásahoch potlačovaná prirodzenosť, ako aj kde leží hranica medzi prirodzeným a umelým, teda antropogénnym, keďže aj človek je v konečnom dôsledku len jeden z prírodných druhov biosféry, existujú isté pragmatické, konvenčne stanovené hranice. Všeobecná zhoda panuje v tom, že ak trváme na kritériu pôvodnosti ako absencii akéhokoľvek ľudského vplyvu, potom pôvodné ekosystémy v súčasnosti prakticky neexistujú, nakoľko minimálne antropogénnymi zmenami atmosféry je priamo či nepriamo ovplyvnená celá biosféra. Naviac, antropogénne ovplyvnený je aj globálny kolobeh vody. Ak však antropický vplyv neponímame takto fundamentalisticky, potom je možné hovoriť o zvyškoch pôvodných ekosystémov v regiónoch, ktoré neboli nikdy v histórii ľudstva súčasťou ekumény, ktoré teda človek priamo nikdy neovplyvňoval, a to ani extenzívnym využívaním. Potom je možné hovoriť napríklad o zvyškoch pôvodných lesných ekosystémov v Južnej Amerike či na Sibíri (pralesoch), kým v Európe ide v najlepšom prípade o pralesovité porasty, ktoré sa vyvinuli po ukončení ľudských zásahov spontánne ako prirodzené. Ako kritérium takejto prirodzenosti možno použiť práve zachovanie základných funkčných väzieb ekosystému, ktoré zaručujú jeho spontánnu obnovu po ukončení antropického tlaku. Takéto prirodzené ekosystémy sa prirodzeným vývojom znova približujú ku kvalite prírodných, z ktorých sú odvodené, možno ich teda označiť aj ako „blízke prírodným“, nie však ako pôvodné. V prípade rieky a jej ohrádzovaného záplavového územia dochádza bez ohľadu na ďalšie hospodárenie či nevyužívanie tohto územia k zásadnej zmene hydrologického režimu. Vplyv záplav sa z väčšiny pôvodnej plochy odstránil, množstvo vody však ostáva rovnaké, a preto je dynamika vodného režimu rádovo intenzívnejšia. Ohrádzované záplavové územie má potom nepôvodnú ekologickú valenciu, tak v hydrologických ako pedologických (a najmä pedogenetických) parametroch. Táto zmenená prirodzenosť vedie k vytvoreniu odvodeného ekosystému, ktorý je odlišný od pôvodného a ponechaný sám na seba neobnovuje kvalitu pôvodnú, ale adaptívnu. Jeho vývoj je totiž usmerňovaný obmedzením, ktoré predstavuje udržiavaná protipovodňová ochrana priľahlého územia. Antropické zásahy do ekosystémov vedú obvykle k vytvoreniu ostrovčekov prirodzených ekosystémov, ktorých ďalší vývoj a prežívanie sú pri výraznom obmedzení pôvodnej plochy silne ovplyvňované exogénne. V prípade rieky a záplavového územia dochádza k inverzii, kedy prvotný exogénny (antropický) zásah vedie k trvalej podpore endogénneho faktora, ktorý sa po silnej redukcii pôvodnej plochy („spútaní“) stáva pre existenciu ekosystému dominantným! V ekosystéme ohrádzovaného záplavového územia sa posilňuje selekcia organizmov smerom k stenotopnosti, viazanej práve na tento typ hydrologického režimu a tvorby pôd. Ak teoreticky pripustíme návrat do stavu bez protipovodňových hrádzí, neprišlo by k samoobnovovaniu ekosystému, ktorý býva v záplavovom území po niekoľkých storočiach existencie hrádzí považovaný za blízky prírodnému či pôvodnému. Naopak ak, znovu teoreticky, pripustíme stav, keď tento dominantný faktor v území vylúčime, akokoľvek benevolentne chápaná prirodzenosť ekosystému sa stráca a jeho prežitie sa stáva závislým na lesohospodárskej pestovateľskej činnosti. Z lesa prirodzeného sa stáva les umelý, neschopný samoobnovy. Tieto úvahy sa týkajú nielen terestrických, ale aj veľkej časti semiakvatických spoločenstiev. Naproti tomu vodné spoločenstvá možno v regionálnom pohľade považovať za pôvodné, len čiastočne narušené prenikaním, alebo vysadením viacerých nepôvodných druhov živočíchov. Zmeny všetkých štyroch typov potamonu sa týkajú podielu ich zastúpenia vo vodnej biote vyplývajúcich z historických, najmä však recentných antropogénnych hydrogeomorfologických zmien. Súčasná miera odvodenosti a nepôvodnosti prírodného prostredia v predmetnom území je výsledkom predovšetkým lesníckych, plavebných, vodohospodárskych ale aj poľnohospodárskych a ďalších antropických zásahov. Čím viac ideme do minulosti, tým je prírodnosť ekosystému vyššia a pre oblasť inundácie typickejšia. Z hľadiska funkčnosti dnešnej inundácie nemôžeme sa vrátiť dozadu pred obdobie, kedy boli postavené protipovodňové hrádze. Pre ďalšie porovnávanie a úvahy o revitalizácii preto považujeme za vhodné približne 50. roky 20. storočia. Rieka Dunaj so svojim ramenným systémom za ostatné storočia prekonala veľké zmeny. Splavné ramená pôvodného toku boli opakovane zanášané hrubými vrstvami naplavenín, čím sťažovali lodnú dopravu. Vtedajší užívatelia dunajskej plavby sa preto začiatkom 19. storočia rozhodli pôvodné plavebné koryto odkloniť doprava, čiže približne do terajšej trasy hlavného toku. Plavbe sa tým pomohlo, ale zároveň došlo k zániku bohatej ramennej sústavy okolo terajšej trasy Malého Dunaja. Okrem toho nové prietočné rameno – teraz nové koryto rieky – odvádzalo hlavné masy vôd do územia, ktoré už predtým predstavovalo skultivovanú poľnohospodársku krajinu. Masy vôd okolo nového toku a jeho ramien zlikvidovali provizorné ochranné hrádze, pustošili sídliská, ničili úrodu, strhávali cesty a zamokrovali pôdu. Sústredená výstavba ochranných hrádzí po obidvoch brehoch nového toku, ktorá bola dokončená koncom 19. storočia, mala stlmiť ničivú silu vôd roztekajúcej sa po celej južnej polovici Žitného ostrova, ale aj Szigetközu. Vegetácia lužných ekosystémov je zákonite viazaná na hydropedologické podmienky ovplyvnené riekou, v tomto prípade Dunajom. Platí to o všetkých typoch rastlinstva, t.j. od fytocenóz vyslovene vodných cez močiarne a brehové typy až po kriačinnú a lesnú vegetáciu. Je to zároveň vegetácia veľmi dynamická, ktorá v porovnaní s klimazonálnymi typmi rastlinstva sa dokáže v priebehu relatívne krátkeho obdobia prispôsobiť meniacim sa podmienkam vodného režimu a sukcesívne vytvoriť stabilné ekosystémy. Odklon a napriamenie rieky a výstavba ochranných hrádzi podstatne zasiahli do pôvodného režimu vôd Dunaja. Zároveň však mali nedozierne následky pre preformovanie sa rastlinstva. Odstavenie vôd pretekajúcich okolo Malého Dunaja podnietilo zarastanie mnohých mŕtvych ramien, čo vyústilo do vzniku zaujímavých spoločenstiev hydro-hygrofytov. Na ich floristické bohatstvo, i keď už značne antropicky pozmenené, poukázal Hejný (1960). Žiaľ, mnohé z nich rozsiahlymi odvodňovacími prácami Žitného ostrova koncom päťdesiatych rokov 20. storočia zanikli. K druhým významným zmenám došlo v medzihrádzovom priestore nového toku Dunaja a jeho ramien, čiže i v terajšom záujmovom inundačnom území. Záplavy sa tu znásobili a búrlivosť ich vôd narástla. Pretrvávanie takého stavu po niekoľko desaťročí začalo selektívne pôsobiť na celé rastlinstvo, preukazne však na lužné dreviny. Tvrdé listnáče ako je dub letný (Quercus robur), menej i jaseň úzkolistý (Fraxinus angustifolia) častú prítomnosť takýchto vôd neznášali a z inundačných území medzihrádzového priestoru postupne vymizli. Na ich miesto nastúpili vŕby a to predovšetkým vŕba biela (Salix alba), menej i vŕba krehká (Salix fragilis) a topole (Populus nigra, Populus alba, Populus x canescens). Vŕbovo-topoľové lesy tu existovali aj predtým, avšak v podstatne menšej rozlohe. Zmena rastlinstva sa dotkla aj nelesnej a vodnej vegetácie. Niekdajšie úseky mŕtvych ramien sa v inundačnom území s častejšími a mohutnejšími záplavami stali prietočnými (eupotamon) a nedovoľovali vznik a existenciu rastlinstva stojatých vôd. Vodná vegetácia mala zabezpečené existenčné podmienky len v ramenách, ktoré boli iba občas prietočné (plesiopotamal) – Oťaheľová (2001). Jej rozvoj ale rapídne začal vo vnútri nížiny a v ramenách odstavených ochrannými hrádzami (paleopotamon). Vývoj brehovej (litorálnej) vegetácie nadväzujúcej na stojaté vody tu bol plošne výraznejší, ale s mnohými prvkami sezónnosti, vyplývajúcimi z kolísania hladín v ramenách (Kubalová 2001). K takýmto pomerom inundácie sa lužné ekosystémy v priebehu 50. až 70. rokov dokonale prispôsobili. Preto prvú komplexnú informáciu venovanú pôdne ekologickým pomerom a lužným lesom Podunajskej nížiny (Jurko 1958) možno považovať za takú, ktorá charakterizuje približne 50. roky 20. storočia, kedy ešte stav narušenia „prirodzeného“ prostredia nebol tak intenzívny. Pri posudzovaní ďalších zmien tento stav vo väčšine prípadov považujeme za východiskový, predstavujúci vegetáciu blízku prírodnej (nie však pôvodnú v pravom slova zmysle) . Žiaľ o vodných a močiarnych spoločenstvách inundácií z tohto obdobia máme len strohé poznámky (Jurko 1958). K ďalším zmenám vodného režimu došlo po rozsiahlych úpravách hlavného koryta rieky spojeného s plavbou a aj s protipovodňovou ochranou v 70. rokoch. Bagrovaním štrku z dna Dunaja na slovenskej i maďarskej strane malo za následok pokles hladiny vody v rieke, a aj hladín podzemných vôd. Tendencia klesajúcej hladiny vody bola neustále udržiavaná pokračujúcou eróziou dna rieky aj pričinením vodných stavieb na rakúskom a nemeckom úseku Dunaja a bagrovania v Dunaji nad Bratislavou. Pokles hladín podzemných vôd ešte pred výstavbou VD bol citeľný zvlášť v hornej časti Žitného ostrova hneď pod Bratislavou, najmä v biskupickom regióne, smerom k Čunovu klesal, ale citeľný bol aj v terajšom inundačnom území. Keďže v inundácii sa už v rokoch úprav koryta Dunaja rozkladali rozsiahle monokultúry šľachtených euroamerických, ale aj našich topoľov, dopad poklesu podzemných vôd tu bol na lesy a ich spoločenstvá menej signifikantný. Už vtedy (dávno pred výstavbou vodného diela) však došlo k poklesu prírastku lesných drevín v úzkom pobrežnom páse Dunaja, najmä na štrkových brehoch (drenážny efekt – Šomšák et al. 1995). Citeľnejší dopad mala úprava koryta na vodné močiarne typy vegetácie ramennej sústavy inundačného územia. Väčšina ramien bola prietočná len pri zvýšených stavoch vody. Nepravidelnosťou prietokov v ramenách však došlo k plošným zmenám medzi spoločenstvami typu plesiopotamon, parapotamon a eupotamon. Podľa osobných skúseností z tohto obdobia tu prevládali spoločenstvá typu plesiopotamon. Tento predpoklad potvrdzujú i novšie výskumy (Oťahelová 2001, Kubalová 2001, Svobodová 1994, Matis 2001). Pripravovaná výstavba Sústavy vodných diel Gabčíkovo – Nagymaros si vyžiadala podrobný floristický výskum celého Podunajska. Touto inventarizáciou tu bolo zistených 959 taxónov cievnatých rastlín. Rozbor viazanosti na stanovištné (fytocenotické) skupiny ukazuje (Šomšák 1999), že z tohto počtu len jedna tretina (311 taxónov) je takých, ktorých život limitujú podzemné a záplavové vody. Sú to vodné a močiarne rastliny (97 druhov), brehové populácie (litorálna, limózna a terestrická ekofáza) so 70 druhmi a nakoniec rastliny, ktorých životný cyklus je viazaný na lužné lesy a kriačiny (194 taxónov). Medzi ostatnými je však obrovský podiel takých druhov, ktoré dokážu a v skutočnosti aj existujú i vo fytocenózach mimo aluviálnych nív (Urtica, Glechoma, Alliaria, Symphytum, Rubus, Poa, Viola, Gagea, Sambucus, Lythrum, Lysimachia a mnohé iné). Ostatné druhy známe zo spomínanej inventarizácie sa viažu na také stanovištia, ktoré nie sú a ani neboli ovplyvňované vodami Dunaja. Sú to napr. druhy xerotermných štrkov (180 taxónov), populácie ruderálnych stanovíšť (190 populácií), obilnín a okopanín (89 taxónov), introdukované druhy (72 taxónov) a neofytné populácie (43 druhov). Stručne povedané až 68,7 % zistených druhov tu existuje bez závislosti na vodách Dunaja (Šomšák 1999). Z ekosozologického hľadiska je však najvýznamnejších práve zvyšných 31%. Popri analýze fytogenofondu boli navrhnuté i určité opatrenia tak, aby výstavbou vodného diela došlo len k minimálnym stratám (Bertová et al., 1986). Na prvom mieste išlo o rozšírenie počtu maloplošných chránených území, ktoré popri existujúcich prírodných rezerváciách budú chrániť celkové stanovištné podmienky vrátane tej vegetácie, ktorej súčasťou sú príslušné druhové populácie. Na úseku Bratislava – Hrušov bolo navrhnutých 9 maloplošných chránených území (Bertová et al., 1986), v terajšej inundácii 7 (Dunajská Sihoť, Šulianské ramená, Kráľovská lúka, Bodícke ramená, Bačianske ramená pri Bake, Istragov a Riečina (Erčéd). Pre sledovanie osudu flóry po výstavbe VD v úseku inundácie boli vyzdvihnuté dva druhy, a to bleduľa letná (Leucojum aestivum) a kotvica kužeľoplodá (Trapa conocarpa). Posledný predstavuje nový druh pre flóru Slovenska a jeho výskyt bol zaznamenaný v mŕtvom ramene pri Kráľovskej lúke /Bodíky (Foto 55). Napriek týmto teoretickým opatreniam v priebehu prípravných prác pred výstavbou VD došlo k citeľnému zásahu do genofondu rastlín, a to na prvom mieste k zmenšeniu areálu mnohých populácií zabratím pôdneho (lesného i nelesného) fondu pre Hrušovskú zdrž, derivačný a odtokový kanál, priesakové kanály a i., (Foto 47). Údaje o flóre tohoto územia sa tesne pred plánovaným uvedením vodného diela Gabčíkovo do prevádzky spresnili podrobnou inventarizáciou spojenou so zakladaním monitorovacích plôch. Uherčíková (2001) ich uvádza v počte 760 taxónov. Ako uvádza autorka už pred spustením prevádzky VD Gabčíkovo tu chýbali viaceré taxóny vzácnych a ohrozených rastlín, ktoré tu boli zaznamenané v 50. rokoch 20. storočia (Hottonia palustris, Gratiola offinalis, Senecio paludosus, Veronica catenata, Sagittaria sagittifolia). Zachovali sa tu však lokality s početným výskytom úplne chráneného druhu Leucojum aestivum. Lesné hospodárstvo realizuje svoje zámery v inundačnej oblasti na rozlohe okolo 3100 ha lesa. Táto rozloha bola od 60-tych rokov len nepatrne pozmenená, a to dokonca jej zväčšovaním. Od 60. rokov dochádzalo k zakladaniu veľkoplošných monokultúr do vopred pripravenej pôdy. V mnohých prípadoch sa zalesnili aj bývalé mŕtve ramená, do ktorých sa počas vytláčania pňov a iných pozostatkov po ťažbe dreva nahrnula skrývka. K hlavným drevinám ako je vŕba biela, vŕba krehká, topoľ čierny, topoľ biely, topoľ sivý sa už v 60. rokoch pridávali kultúry cudzokrajných topoľov. Už okolo roku 1956 sa ich rozloha pohybovala okolo 27 % z existujúcej rozlohy lesov (Jurko 1958). Od roku 1956 sa ich plošný podiel prudko zvyšoval a už okolo roku 1981 dosahovala v dunajských lužných lesoch okolo 80 % (Vojtuš 1986). V prvých začiatkoch to boli kultivary Populus deltoides – „monilifera“ a Populus x euroamericana – „robusta“ a neskôr i rajonizovaný klon „I-214“ vyšľachtený v Taliansku (Neštický, Varga 2001). Lesnícky výskumný ústav Zvolen – Výskumná stanica Gabčíkovo už od roku 1956 zakladal v tomto území pokusné plochy. Ich cieľom však bolo overovanie lesopestovateľských zásahov do monokultúr šľachtených drevín (východiskový spon pri zalesňovaní, intenzita výchovných zásahov) a nie ekologický monitoring. Zo získaných poznatkov rozmanitých klonov topoľov a vŕb od začiatkov ich cieleného pestovania až do prehradenia Dunaja sa však potvrdil predpoklad, že ide o dreviny a porasty tolerantné voči menším zmenám vodného režimu (Varga 1993, Neštický, Varga 2001). Za pestovanie euroamerických topoľov a ich kultivarov sa prihovárala aj nezvyčajne vysoká produkcia drevnej hmoty. Prvé merania signalizovali ročný prírastok až 25 m3/ha. Pokles prírastku drevnej hmoty signalizovali len porasty ľavobrežného pásu Dunaja medzi Čunovom a Sapom. Dendroekologické bádania tohto úseku však potvrdili pokles prírastkov dávno pred prehradením Dunaja (bagrovanie dna rieky – Šomšák et al., 1995). Na základe dostupných literárnych prameňov, ale aj z autentických poznatkov Šomšáka z týchto rokov, možno stav inundačného územia Dobrohošť – Sap pred prehradením Dunaja charakterizovať nasledovne:
Terestrické živočíšne spoločenstvá inundačnej oblasti (ale aj ďalších častí Podunajska snáď s výnimkou xerotermných) sa v minulosti študovali len sporadicky a o väčšine z nich chýbajú údaje, ktoré by umožňovali načrtnúť stav pôvodných zoocenóz dotknutého územia. Väčšina prác z obdobia pred monitoringom je orientovaná faunisticky, prípadne na základné ekologické otázky, napríklad väzbu na pôvodné, neraz už pozmenené alebo zaniknuté biotopy (Štepanovičová 1995). Pri definovaní pôvodného stavu spoločenstiev terestrických živočíchov záujmového územia Podunajska možno vychádzať len čiastočne z údajov získaných priamo z tejto oblasti pred uvedením VD Gabčíkovo do prevádzky, ich informačná hodnota je však obmedzená, pretože aj vtedajší stav bol výsledkom vyše storočného radikálneho zasahovania do hydrologického režimu aluviálnych ekosystémov. Konzistentné dáta boli získané až od začiatku monitoringu bioty v roku 1990, v súvislosti s výstavbou a prevádzkou vodného diela Gabčíkovo (Jedlička et al., 1999). Za relatívne pôvodný stav môžeme považovať zoocenózy zvyškov lužných lesov v medzihrádzovom priestore v úseku Sap – Čičov (Foto 62, Foto 63), ktoré bývajú aj v súčasnosti zaplavované a zásobované priesakovou vodou, a štruktúrou sa najviac podobajú pôvodným spoločenstvám najvlhších typov mäkkých lužných lesov, súdiac podľa prevahy polyhygrofilných druhov adaptovaných na podmienky cyklického klimaxu. Pri rekonštrukcii východiskového stavu či zloženia taxocenóz pred prehradením Dunaja a odvedením vôd do derivačného kanála koncom roka 1992 je možné sa čiastočne opierať aj o údaje o zoocenózach z blízkych lokalít alúvií riek Moravy, Malého Dunaja a Váhu. Uvedené materiály môžu slúžiť tak na definovanie prírodného stavu ako aj jednotlivých stavov spoločenstva, viac alebo menej blízkych prírodným. Terestrická fauna a zoocenózy v inundácii sú typické pre vnútrozemskú deltu Dunaja s bohatou sieťou riečnych ramien a stojatých vôd súvisiacou s ukladaním transportovaných častí a meandrovaním rieky a častými záplavami. Tento dynamický systém je špecifickým komplexom ekosystémov vodného, pôdneho a suchozemského prostredia a tomu zodpovedajúcich ekotonov, v terestrickej časti Phragmition, Magnocaricion elatae, Caricion gracilis, Oenanthion aquaticae, Elatino-Eleocharition ovatae, Chelidonio-Robinion, Lolio-Potentillion, Salicion albae, Ulmenion, Asparago-Crataegetum. Jednotlivé spoločenstvá predstavujú katenu podĺž vlhkostného gradientu súvisiaceho s výškou a kolísaním hladiny podzemných vôd a existenciou sezónnych záplav. Celý systém patrí k typu cyklických klimaxov udržiavaných cyklickým hydrologickým režimom rieky a celého územia. Zmeny hladiny podzemných vôd a záplav s následnými zmenami vlhkostného gradientu môžu vyvolať zmeny kateny od jednoduchých posunov v priestore až po posun ku xerickým spoločenstvám a vymiznutie hygrofilných spoločenstiev. Dominantným faktorom ovplyvňujúcim prirodzené spoločenstvá je teda hladina podzemnej vody súvisiaca s infiltráciou z hlavného koryta. Špecifické podmienky ekosystému podmieňujú existenciu špecializovaných druhov adaptovaných morfofyziologicky, životnými cyklami a stratégiou na prežívanie v podmienkach cyklického klimaxu. V zoocenózach sa podstatnou mierou prejavujú druhy ekologicky špecializované, ktorých existencia mimo týchto podmienok spravidla nie je možná. Rovnako je však ohrozená aj v situácii, ak zmeny podmienok sú síce v hraniciach ich ekologickej tolerancie, avšak umožňujú prenikanie široko tolerantných druhov, ktorým špecializované druhy nie sú schopné konkurovať. Existencia vysoko špecializovaných druhov sa prejavuje v sformovaní špecifických zoocenóz, udržiavaných a podmienených práve podmienkami vnútrozemskej delty s cyklickými katastrofami. V plochých nížinných alúviách sa vytvárali charakteristické spoločenstvá s vysokými nárokmi na vlhkosť, ktoré vďaka šírke nivy neboli v relatívne širokom pásme infiltrované mezohygrofilnými druhmi. K vzájomnému prelínaniu druhov dochádzalo až vo väčšej vzdialenosti od toku, prípadne až na samotnom okraji nivy. Šírku tohto prechodného pásma určovala konfigurácia terénu a vzájomný kompetičný tlak druhov oboch ekologických skupín. Poloha tohto pásma však nebola stála, ale dynamicky sa menila podľa výkyvov vodnatosti toku. Táto situácia umožňovala, podľa zmien polohy koryta, rozsahu záplav a výšky hladiny podzemnej vody na jednom mieste rýchle alternovanie rôznych, ale vždy prírodných spoločenstiev. Ide o situáciu, aká bola v dávnej minulosti typická pre rozsiahle nížinné územia, teda aj pre oblasť širšieho okolia VD Gabčíkovo. Formovanie tohto prirodzeného gradientu spoločenstiev v záujmovej oblasti obmedzujú protipovodňové hrádze, ktoré odrezali od prirodzeného hydrologického režimu časť lesných porastov v mimohrádzovom priestore. V dôsledku toho sukcesia spoločenstiev v mimohrádzovom priestore má predpoklady smerovať k mezohygrofilným spoločenstvám lesných geobiocenóz normálneho hygrického radu, prípadne pri narušení integrity porastu k spoločenstvám nelesných ekosystémov. Naopak časť spoločenstiev v užšom medzihrádzovom priestore je pravdepodobne vystavená intenzívnejšiemu mechanickému pôsobeniu záplav usmernených do pomerne úzkeho koridoru a zbavených možnosti rozliať sa do odľahlejších časti alúvia. Jeden i druhý stav sa teda odlišuje aspoň na časti sledovaného územia od stavu, ktorý tu panoval v prírodnej, alebo človekom len zanedbateľne zmenenej krajine. Zoocenózy ako spoločenstvá konzumentov a reducentov sú v celom sledovanom území viazané na (Jedlička et al., 1999):
Taxocenózami, ktoré veľmi dobre dokumentujú nielen stav ale aj vývoj zoocenóz, sú malakocenózy. Je známe, že taxocenózy mäkkýšov (Mollusca) prešli v kvartéri zložitým sekulárnym vývojom (Ložek 1955), súčasné spoločenstvá sa formovali postupne v mladšom holocéne (cf. Cheben a kol. 2001) avšak pravdepodobne na omnoho rozsiahlejšom území, takže dnešný pás v medzihrádzovom priestore je len ich zvyškom. Podľa doterajších nálezov holocénnej malakofauny je pravdepodobné, že približne v oblasti dnešnej obce Vrakúň kedysi prebiehala hranica oddeľujúca široký pás zaplavovaných luhov od vyvýšeného, suchšieho, čiastočne stepného vnútra Žitného ostrova. Inundačné územie na dnešnej slovenskej strane malo teda v minulosti (mladšom holocéne) šírku približne 10 km (dnes sú to v najširšom mieste pri Bake 3 km) (Ložek 1955). Otázke stepných enkláv sa venuje v rámci záplavového územia Dunaja najmä Ložek (1955, 1964). Zatiaľ chýba časové datovanie jednotlivých dejov aj znalosti o topografickom rozložení jednotlivých geobiocenóz, čo môže upresniť len ďalší a širší výskum recentnej a fosílnej malakofauny, pravekých sídlisk a dopĺňanie znalostí z archívnych údajov tejto oblasti. Analýzy tanatocenóz sedimentov Dunaja vo Vlčom Hrdle pri Bratislave kombinované s absolútnym datovaním veku sedimentov (Pišút 2000, Čejka 2000) umožnili zrekonštruovať kompletnú sukcesnú líniu lužného lesa od iniciálneho štádia lužného lesa až po dnešný prechodný lužný les (obdobie r. 1791-1999, t.j. 208 rokov) a potvrdili výskyt podobných typov lužného lesa, aké sa na Podunajsku vyskytujú v súčasnosti a tanatocenózy, ktoré štruktúrou zodpovedajú približne dnešným zvyškom najzachovalejších lesných malakocenóz v oblasti medzi obcami Dobrohošť a Sap. Celková plocha najvlhších typov mäkkých lužných lesov s príslušnými malakocenózami bola v minulosti oveľa väčšia než dnes, s prirodzenejšou štruktúrou porastov, čo je zreteľne vidieť dokonca už pri porovnaní fytocenologických zápisov zo 40-50-tych rokov so súčasnými (Uherčíková 1995, 1998). Charakteristické hlavne pre iniciálne štádiá tzv. mäkkého luhu a iné stanovištia s vysokou pôdnou vlhkosťou sú najmä výrazne vlhkomilné druhy ulitníkov Succinea putris, Oxyloma elegans, Zonitoides nitidus a Pseudotrichia rubiginosa. Diferenciačnými druhmi vlhkých typov mäkkého lužného lesa (asoc. Salici-Populetum myosotidetosum až Salici-Populetum typicum Jurko 1958) sú, okrem vyššie uvedených druhov aj polyhygrofilný Carychium minimum a lesné hygrofilné druhy Arianta arbustorum, Vitrea crystallina a sčasti aj Urticicola umbrosus. Pre tzv. prechodný až tvrdý luh (as. Fraxino-Populetum, Fraxno-Ulmetum) je zase typická dominancia prevažne lesných mezohygrofilných druhov, ktoré neznášajú ničivý vplyv záplav a dlhodobo podmáčanú pôdu (Aegopinella nitens, Cochlodina laminata, Semilimax semilimax, Alinda biplicata, Monachoides incarnatus, Petasina unidentata, Clausilia pumila, čiastočne aj Carychium tridentatum). V taxocenózach sú zastúpené aj skupiny druhov, ktoré sú viazané na vyslovene nelesné stanovištia, alebo riedko zapojené porasty stromov či krov (Vallonia pulchella, V. costata, Euomphalia strigella, Cepaea vindobonensis a Xerolenta obvia). Analogicky je možné predpokladať podobné pomery (pokiaľ ide o zastúpenie jednotlivých ekoelementov najmä vo vzťahu k vlhkosti, vode, zatieneniu atd.) aj v iných taxocenózach terestrických živočíchov. Vo faune suchozemských rovnakonožcov (Oniscidea) z Podunajska v dosahu vodného diela Gabčíkovo bolo z obdobia 1986-1990 zistených 16 druhov (Flasarová 1999), najpočetnejším bol eurytopný Trachelipus rathkei. Stonožkám (Chilopoda) Podunajska bola v období pred zahájením výstavby VD Gabčíkovo venovaná len okrajová pozornosť a podobne ako u predchádzajúcej skupiny netýkajú sa priamo územia tejto štúdie, ale jeho okolia (Gulička 1957, Čarnogurský et al., 1994). Za východiskový stav taxocenóz stonožiek sa môže do istej miery považovať druhové zloženie zistené v úseku derivácie pred rokom 1993, na monitorovacích plochách bolo zaznamenaných 9 – 14 druhov. Získané údaje o spoločenstvách suchozemských rovnakonožcov (Oniscidea) a stonožiek (Chilopoda) sú veľmi blízke až totožné s údajmi z lesov alúvií Moravy a Dyje na moravskej i rakúskej strane (Tajovský 1999, Tuf 2000, Zulka 1999), môžeme ich považovať za dostatočne charakteristické. V Dobrohošti - Dunajských krivinách z 13 zistených druhov k eudominantom patril eurytopný Lithobius forficatus (D = 21%-23%), z okolia sem prenikal Lithobius mutabilis. Z pravidelne sa vyskytujúcich druhov, ktoré sú indikátormi vyššieho stupňa pôdnej vlhkosti (od 20%-30%) boli zistené mezohygrofilné Lithobius curtipes, L. crassipes, Geophilus flavus. Lithobius microps tu už po roku 1991 zistený nebol. Spoločenstvo stonožiek Bodíckej brány tvorilo 13 druhov, medzi ktoré patrili mezohygrofilné L. crassipes a L. curtipes s vyššími hodnotami dominancie (17%-18%) a hygrofilný L. agilis, náročný na pomerne vysoký stupeň pôdnej vlhkosti. Tretinu druhov však predstavovali eurytopné druhy L. forficatus, L. erythrocephalus a L. mutabilis, ktoré boli a sú zastúpené po celom sledovanom území Podunajska. Taxocenózu stonožiek Kráľovskej lúky tvorilo pred prehradením 14 druhov, ku ktorým patrili typický ripikolný a hygrofilný Lamyctes emarginatus a hygrofilné druhy L. agilis a L. microps. Eudominantným druhom bol Lamyctes emarginatus, dominantnými Lithobius aeruginosus a L. curtipes. Spoločenstvo stonožiek Kráľovskej lúky pred prehradením Dunaja zaraďujeme medzi najbohatšie. Za obdobie rokov 1991 – 1997 tu bolo zistených 17 druhov. V prvom roku po prehradení (1993) počet zaznamenaných druhov bol zreteľne nižší oproti rokom predchádzajúcim. Absentujúce druhy sa opäť objavili až v roku 1997, naviac v tomto roku bol po prvýkrát zistený Lithobius pelidnus, druh s centrom rozšírenia vo východnej Európe preferujúci zalesnené územia. V spoločenstve stonožiek Istragova bolo zastúpených 9 druhov, z ktorých medzi eudominantné patrili eurytopné Lithobius forficatus a L. mutabilis. Z typických mezohygrofilných druhov boli vysokým percentom zastúpené tiež L. aeruginosus, L. crassipes, L. curtipes a hypogeicky žijúci druh Pachymerium ferrugineum. Pred prehradením Dunaja nechýbal ani hygrofilný druh L. agilis zistený v rokoch 1991 a 1992. Z územia, na ktorom je v súčasnosti vybudované VD Gabčíkovo a jeho najbližšieho okolia viacerí autori (Brtek, Rotschein 1964, Šiška 1983, Štepanovičová, Lapková 1984, Bulánková 1995, Štepanovičová 1989, 1991) zistili pred začatím výstavby výskyt 177 druhov bzdôch (Heteroptera). V epigeickej taxocenóze bystruškovitých (Coleoptera: Carabidae) v podunajských lužných lesoch s pravidelnými záplavami prevládali s vysokou dominanciou vlhkomilné druhy. Na Dunajských krivinách to boli Agonum moestum (13,35%), Oxypselaphus obscurus (22,39%), Patrobus atrorufus (11,87%) a Platynus assimilis (3,56%). Jednotlivo sa objavovali aj ďalšie vlhkomilné alebo ripikolné druhy (napr. Bembidion dentellum, Bembidion biguttatum, Oodes helopioides). Podobne v Bodíckej bráne výrazne prevažovali vlhkomilné druhy P. assimilis (12,96%), P. atrorufus (12,41%), O. obscurus (3,31%) sprevádzané ďalšími početne zastúpenými vlhkostne menej náročnejšími, resp. tolerantnejšími druhmi Asaphidion flavipes (30,42%), Carabus granulatus (8,09%), Pterostichus strenuus (9,65%), Clivina fossor (4,32%). Významná bola aj prítomnosť vlhkomilných druhov B. dentellum a Badister sodalis. Na Kráľovskej lúke, kde bolo spoločenstvo sledované už od roku 1987, prevažovali paludikolné A. moestum (15,95%), vlhkomilné druhy P. atrorufus (13,79%), P. assimilis (4,96%) a relatívne tolerantné druhy P. strenuus (29,31%), C. granulatus (7,76%) a O. obscurus (4,31%). Vysokej vlhkosti, resp. podmáčaniu zodpovedala prítomnosť Europhilus fuliginosus, Europhilus micans, Oodes helopioides. V roku 1989 sa abundancia všetkých vlhkomilných druhov okrem A. moestum zvýšila. Veľmi prudko vzrástlo zastúpenie A. flavipes (z 2 na 248 jedincov). V rokoch 1990-1991 bol celkový úlovok v dôsledku zaplavenia lokality nižší (z 1348 jedincov v r. 1989 klesol na 727, resp. 372 jedincov), ale štruktúra spoločenstva sa výrazne priblížila pôvodnému stavu z roku 1987 (proporcionálna podobnosť 56,7-57,6%, podobnosť abundancie 39,6-40,5%). Zaplavenie postihlo aj dva vlhkomilné, ale nie vyslovene paludikolné druhy P. atrorufus a P. assimile. Na Istragove v roku 1989 výrazne prevažovali dva drobné vlhkomilné druhy bystruškovitých A. flavipes (47,7%) a Bembidion femoratum (15,36%), výrazné zastúpenie mali aj ďalšie vlhkomilné druhy P. assimile (15,5%), P. strenuus (5,7%), C. granulatus (3,6%), P. atrorufus (2,4%). Ekologicky významná bola prítomnosť druhov Europhilus fuliginosus a E. micans. Vysoké zastúpenie vlhkostne menej náročných druhov ešte pred podstatnými zmenami hydrologického režimu odráža prevahu silne piesčitej pôdy na veľkej časti sledovanej plochy. Načrtnutý stav naznačuje, že taxocenózy bystruškovitých predstavovali charakteristické spoločenstvá druhov s vysokými nárokmi na vlhkosť (Tab. 2.1, 2.2), ktoré neboli v podstatnej miere infiltrované mezohygrofilnými druhmi obývajúcimi geobiocenózy normálneho hydrického radu. Neboli ani výrazne ovplyvnené druhovou skladbou porastov, ktorá bola v ostatných desaťročiach silne narušená náhradou pôvodných porastov topoľovými kultúrami. Príčinou je skutočnosť, že pre hygrofilné druhy lužných lesov je rozhodujúca prítomnosť v podstate akéhokoľvek hustého vegetačného krytu, ktorý zabraňuje vysušovaniu a prehrievaniu hrabanky priamym slnečným žiarením. Takéto podmienky boli schopné pre bystruškovité vytvárať tak topoľové kultúry staršie ako 10 rokov, t. j. počas prevažnej časti svojej existencie ako aj vysoký bylinný porast, ktorý sa (bez ohľadu na svoje floristicky nevyhovujúce druhové zloženie takýchto porastov) vyvinul na rúbaniskách v podmienkach dostatočného zásobenia pôdy vlhkosťou. Pre semiakvatické, amfibické a prechodné živočíšne taxocenózy je pomerne charakteristickým javom ich väzba nielen na vegetáciu ako potravnú bázu, ale aj viazanosť na vodný režim; jeho nepravidelné zmeny s následnou sukcesiou sa prejavujú na nestabilite zloženia taxocenóz a ich veľkých medziročných zmenách, čo dokumentuje situácia taxocenóz fytofágnych Curculionidae (Coleoptera) brehových vegetačných formácií v systéme dunajských ramien a hlavného toku Dunaja, staršie údaje o ktorých sú v širšie koncipovaných štúdiách (Majzlan, Rychlík 1982, Majzlan 1988, 1990, Kodada, Majzlan 1991) a neskôr boli monitorované. Z uvedených prác vyplýva, že pobrežné územia niektorých skúmaných ramien boli už pred prehradením Dunaja do značnej miery aridizované. Odrazilo sa to aj na pomernom zastúpení eurytopných a stenotopných druhov: hygrofilných a paludikolných na jednej a druhov xerofilných a na biotop nenáročných na strane druhej. Je možné sa oprávnene domnievať, že spoločenstvo nosáčikov (Curculionidea) zistené v rokoch po prehradení Dunaja (1992) žilo na skúmanom území v pobrežnej vegetácii ramien vnútrozemskej delty aj pred jeho prehradením. Z vyschnutých ramien sa táto taxocenóza stiahla do menších enkláv, v ktorých prežívala. S časom prehradenia Dunaja koinciduje šírenie sa na sever hygrofilného a a ripikolného nosáčika Bagous bagdatensis. Jeho lokality na území Slovenska predstavujú dosiaľ známu severnú hranicu rozšírenia. Ripikolný a akvikolný Dicranthus majzlani, na území Slovenska aj v celej Európe kriticky ohrozený, je indikátorom prírodne zachovalých stojatých a polotečúcich nížinných vôd a vyžaduje vyššiu hladinu vody v ramennom systéme. Obidva druhy neboli do roku 1992 z tohto územia známe. Tab. 2.1 Štruktúra prirodzeného spoločenstva bystruškovitých v lužnom lese často zaplavovanom prúdiacou vodou; druhy sú zoradené podľa klesajúcich vlhkostných nárokov a počtu jedincov Carabidae natural structure in the fooodplain forest often flooded by flowing water; species are arranged according to their moisture demand and number of individuals
Tab. 2.2 Spoločenstvo bystruškovitých v silne narušenom zvyšku lužného lesa v blízkosti zvyšku mŕtveho ramena Malého Dunaja Carabidae taxocoenoses in heavy disturbed rest of floodplain forest close to the rest of Malý Dunaj dead arm
Symboly ako v Tab. 2.1 – Symbols as in Table 2.1 V priebehu monitoringu sa zistilo, že zatiaľ čo v komplexe pobrežných rastlinných spoločenstiev (asociácie Rorippo-Agrostietum stoloniferae, Phalaridetum arundinaceae, Rorippo amphibiae-Oenanthetum aquaticae, Glycerietum maximae, Phragmitetum communis, Caricetum gracilis) bolo v taxocenóze Curculionidae zistených 49 druhov, z toho 13 (29%) každoročne (Sitona macularis, Sitona suturalis, Bagous collignensis, B. glabrirostris, Tanysphyrus lemnae, Rhinoncus albicinctus, R. perpendicularis, R. inconspectus, Poophagus sisymbrii, Tapinotus sellatus, Nanophyes brevis, N. globiformis, N. marmoratus) s vyrovnaným pomerom hygrofilné druhy viazané na pobrežnú vegetáciu, ako aj druhov viazaných na rastliny vodnej hladiny, so signifikantnou prevahou charakteristických, stenotopných a hygrofilných druhov, vo vysychajúcom slepom ramene Dunaja v lužnom lese (Salici-Populetum) s asociáciou Phragmitetum communis s väčším množstvom vody iba v jarnom období (apríl a máj) bolo v taxocenóze Curculionidae zistených 39 druhov, pričom ani jeden druh sa nevyskytoval každoročne a eudominantným bol sprievodný druh Nedyus quadrimaculatus. Zo záujmového územia boli dávnejšie pomerne dobre známe taxocenózy suchozemských resp. amfibických stavovcov. Na sledovanom území bol známy výskyt 12 taxónov (11 druhov + 1 klepton) obojživelníkov, z nich Triturus dobrogicus a Rana ridibunda sú v kategórii ohrozených (EN), Triturus vulgaris a Rana lessonae v kategórii zraniteľných (VU), všetky ostatné v kategórii rizikových (LR) druhov. Z 12 druhov plazov známych z územia Slovenska sa tu vyskytuje 9, z toho 7 chránených, 3 v kategórii zraniteľných (VU: Coronella austriaca, Natrix tesselata, Lacerta viridis), ostatné v kategórii rizikových (LR) druhov. Z ornitologického hľadiska predstavovali podunajské lužné lesy spolu s ramenným systémom Dunaja ešte v nedávnej minulosti územie s vysokou diverzitou a denzitou druhov, kde hniezdili viaceré vzácne a ohrozené druhy vtákov (Balát 1963). Hniezdnu ornitocenózu podunajských lužných lesov pred výstavbou VD Gabčíkovo – t. j. v 70. a 80. rokoch – tvorilo 103 druhov vtákov (Tab. 2.3). Z významných hniezdičov to boli predovšetkým haja tmavá (Milvus migrans - VU) a chochlačka bielooká (Aythya nyroca - EN), ktoré tu vytvárali hniezdne populácie celoslovenského významu, ďalej bučiačik močiarny (Ixobrychus minutus VU), bocian čierny (Ciconia nigra), včelár lesný (Pernis apivorus), rybárik riečny (Alcedo atthis), ďateľ prostredný (Dendrocopos medius), ktoré tu vytvárali hniezdne populácie nadregionálneho významu. Z celkového počtu 103 hniezdičov boli 3 druhy ohrozené (EN: Ardea purpurea, Aythya nyroca, Coracias garrulus), a 4 zraniteľné (VU: Ixobrychus minutus, Milvus migrans, Nycticorax nycticorax, Upupa epops). Tab. 2.3 Prehľad vtákov, ktoré hniezdili v podunajských lužných lesoch a v priľahlom ramennom systéme Dunaja (slovenská strana) pred výstavbou VD Gabčíkovo (1970 - 1990) Review of fowls nesting in the Danube floodplain forest and river arm system on the Slovak side before putting the Gabčíkovo project into operation (1970 - 1990)
Vysvetlivky: H – druh hniezdi na protiľahlej, maďarskej strane Dunaja, jeho potravný revír však zasahuje aj na slovenskú stranu h – hniezdil v období tesne pred výstavbou i po sprevádzkovaní VD nzh – nepravidelný zriedkavý hniezdič sh – synantropný hniezdič, preniká popri horárňach a chatových osadách X – druh hniezdil pred výstavbou, po sprevádzkovaní nebolo hniezdenie zistené Okrem lužných lesov dôležitých pre hniezdiče bol Dunaj významnou trasou migrácie vodného vtáctva. Na hlavnom toku Dunaja zimovalo v jednotlivých rokoch 25-30 druhov vtákov (Kalivodová, Darolová 1998, Áč et al., 1996). Medzi dominantných hibernantov patrili Anas platyrhynchos a Bucephala clangula. Vo faune cicavcov (Mammalia) bolo zistených 49 druhov. Bobor (Castor fiber), na území vyhynutý, sa v súčasnosti po reštitúcii v Rakúsku spontánne šíri, nenachádza však v študovanom území optimálne podmienky. V taxocenóze drobných zemných cicavcov, tvorenej druhmi Sorex araneus, Apodemus flavicollis, Clethrionomys glareolus, Sorex minutus, Crocidura leucodon, Crocidura suaveolens, Microtus arvalis, Microtus oeconomus, Pitymys subterraneus, Apodemus sylvaticus, Micromys minutus dochádza v závislosti najmä na vlhkostnom gradiente k zmene štruktúry dominancie. V mäkkom lužnom lese sú eudominantné Sorex araneus, Apodemus flavicollis a Clethrionomys glareolus, s posunom na vlhkostnom gradiente smerom ku xerickým podmienkam sa ich dominancia znižuje a vo zvýšenej miere sa v spoločenstve uplatňujú iné druhy vrátane tu nepôvodných Microtus arvalis a Mus musculus. Štruktúra, dynamika a produkcia planktónu, bentosu, nektónu i neustónu verne odráža pôvodnosť tokov a kvalitu vôd. Spoločenstvá týchto organizmov predstavujú významnú zložku charakterizujúcu tok a jeho ramennú sústavu a sú významnými indikátormi zmien, schopnými odpovedať na zmeny v rôznych časových rovinách a tak sa z ich charakteru môžu vyvodzovať závery o zmenách, ktoré prebehli, prebiehajú a umožňujú aj v základných rysoch predpokladať ďalší vývoj. Technické zásahy, s cieľom znížiť riziká povodní a zlepšiť plavebné podmienky, sa sústredili pôvodne predovšetkým na prehlbovanie a upravovanie dna, regulácie brehov, likvidáciu lužných pralesov, uzatváranie ramien (DUB, SZOLGAY, 1966). Výsledkom toho bola výrazne zvýšená rýchlosť prúdenia v rieke a silná erózia dna, výrazné zníženie a prerušenie prúdenia v pôvodne prietočných ramenách typu eupotamál a parapotamál. Pôvodná heterogenita mezohabitatov bola potlačená. Výrazne sa sťažili podmienky migrácie lariev, ale aj reprodukcie imág vodného hmyzu a iných vodných organizmov (KRNO, 1995). Uzatváraním vtokov ramien pre zabezpečenie plavby a v dôsledku poklesu riečiska Dunaja sa vytvorili predovšetkým podmienky pre rozšírenie ramien typu plesiopotamál charakterizovaných rozvojom makrofýt, eutrofizáciou a ich postupnú premenu na vysýchajúce typy periodických mokradí. Vybudovanie protipovodňových hrádzí spôsobilo stratu spojenia ramien v mimohrádzovom priestore s aktívnym hlavným tokom a ich premenu na ramená typu paleopotamál. Takéto podmienky znamenali ústup reofilných, potamofilných a stenoekných foriem makrozoobentosu a ich nahradenie skôr eurytopnými formami. Negatívne pôsobilo aj postupné rozpojovanie systému siete ramien v priestore medzi protipovodňovými hrádzami. Stössel (1988) a Fruget (1991, 1992) poukázal na vplyvy regulácií na štruktúru makrozoobentosu veľkých európskych riek (Rýn, Rhôna), v ktorých dochádza komplexne k homogenite spoločenstiev a k narušeniu longitudinálnej zonácie. Pre Dunaj a jeho inundáciu boli v minulosti typické viac-menej pravidelne sa opakujúce vyššie prietoky začiatkom jari v marci a začiatkom leta v júni a júli (Mucha et al., 2001). To svedčí o špecifickosti prietokového režimu tejto rieky, a preto ju nemôžeme označiť ako typicky alpský typ (Holčík et al., 1981). Výrazné jarné maximá sú výsledkom topenia snehov v predpolí Álp a Hercynika, takže ide o zmiešaný typ vysokohorského a podhorského prietokového režimu. Takémuto vodnému režimu boli podriadené aj vývinové cykly temporárnej epifauny Dunaja a jeho inundácie. Záplavy, periodické i neperiodické, boli hlavným prírodným faktorom genézy ekosystémov Dunaja (Rovný et al., 1996). Záplavy preplachovali mediál ramien, zabraňovali kolmatácii dna, najmä jarné bohato zásobovali ramená živinami, priaznivo ovplyvňovali kyslíkový i teplotný režim, umožňovali tak úspešný ontogenetický vývin hydrobiontov a v neposlednom rade uľahčovali aj ich pasívnu i aktívnu migráciu. Protipovodňovými hrádzami a smernými stavbami v toku bol tento režim zmenený, nastalo k rýchlejšie a väčšie kolísanie hladín, zrýchlenie rýchlosti toku najmä v prehlbovanom úseku napriameného hlavného koryta, nastal pokles hladín povrchových i podzemných vôd. Od 60. rokov 20. storočia bol tento proces sprevádzaný aj postupným uzatváraním ramien, fragmentáciou vnútrodunajskej delty, poklesom frekvencie preplachovania ramien až ich úplným vyschnutím. Faunistické údaje mikro-meiozoobentosu tejto oblasti boli publikované vo viacerých prácach (Ertl, 1966, 1970). Najviac údajov bolo známych o perifytóne, ktorý tvorili predovšetkým malé meňavky (Amoebina), niektoré druhy nálevníkov a sesilných Rotifera (Matis, Tirjaková (1992, 1995a, 1995b), Tirjaková (1992, 1999), Baláži, Tirjaková (1999), Baláži, Matis (2002)). Makrozoobentos Dunaja, jeho ramien a periodických vôd bol predmetom štúdia viacerých autorov: Balthasara (1936, 1938), Hraběho (1941), Trpiša (1957, 1962), Raušera (1957), Lichardovej (1958), Ertlovej (1963, 1968, 1970, 1973, 1987), Brteka a Rothscheina (1964), Russev (1979), Pomichala (1984), Landu (1969), Nagya a Šporku (1990), Krna (1990), Majzlana (1992), Davida (1994), Bulánkovej (1995, 1999, 2001), Bulánkovej a Halgoša (1995, 1997), Halgoša (1995), Šíbla et al. (2001), Šporku, Krna (1995), Košela (1995a) a Šporku (1980a, 1980b, 1982, 1983, 1984, 1994, 1998 ), Šporku, Nagya (1998). Z týchto prác vyplýva, že Dunaj a priľahlé inundačné územie je už vyše storočia pod silným antropickým tlakom, ktorého výsledkom je napríklad vyhynutie viac než 90% druhov dunajských pošvatiek, viac než 50% podeniek a viac než 30% potočníkov (Krno a kol., 1999). Územie Žitného ostrova bolo od dávna známe kvantitatívnym i kvalitatívnym bohatstvom komárov (neustón), ako i každoročnými kalamitami súvisiacimi s premnožením niektorých druhov. Kalamity spôsobovalo vyliatie vôd z Dunaja do inundačného územia. Trpiš (1962) udáva z územia Žitného ostrova 28 druhov komárov. Od tohto obdobia prešlo územie veľkými zmenami súvisiacimi s výraznými antropickými zásahmi (Halgoš, 1981; Halgoš, 1984). Spoločenstvá zooplanktónu pred výstavbou vodného diela Hlavný tok Dunaja Pred prehradením Dunaja (1971-1972) (Vranovský 1974) a v rokoch 1991-1992 (Illyová 1995; Vranovský, Illyová 1999) dominovali medzi planktonickými vírnikmi hlavného toku spravidla euplanktonické druhy, najmä z rodov Keratella (K. cochlearis), Polyarthra (P. vulgaris, P. remata), Synchaeta (S. oblonga, S. tremula, S. stylata), Brachionus (B. calycilforus, B. angularis), a v zimnom období aj Rotaria rotatoria. V krustáceoplanktóne podobne prevládali pravé planktonty, avšak v hornej časti monitorovaného úseku (na profile Dunajské kriviny) značný podiel z celkovej abundancie pripadal aj na tychoplanktonické (litorálne a bentické) druhy, t.j. nepravé planktonty. Z perloočiek (Cladocera) dominovala Bosmina longirostris, prípadne spolu s druhom Daphnia longispina a/alebo D. cucullata, z veslonôžok (Copepoda) najčastejšie Acanthocyclops robustus, a to buď so vznášavkou Eudiaptomus gracilis alebo s druhmi Cyclops vicinus a Thermocyclops oithonoides, prípadne aj s druhom Eurytemora velox (imigrant, zistený v našom úseku Dunaja prvýkrát v roku 1991). Z tychoplanktonických perloočiek k najhojnejším patrili Alona quadrangularis a A. affinis, z veslonôžok Eucyclops serrulatus . Ramená typu parapotamál Pred prehradením Dunaja vo vodách tohto typu sa v teplej časti roka v obdobiach stagnácie prietočnosti spravidla vyvinul kvantitatívne bohatý zooplanktón tvorený výlučne euplanktonickými druhmi (Vranovský 1974, 1985; Vranovský, Illyová 1999). Z vírnikov dominovali viaceré druhy rodov Brachionus, Keratella, Polyarthra a Synchaeta. Z perloočiek to boli Bosmina longirostris spolu s Daphnia longispina a D. cucullata (v ramenách nad Gabčíkovom), resp. s D. cucullata, Diaphanosoma brachyurum a Moina brachiata (v Istragovskom ramene - pod Gabčíkovom). Podobne z druhej významnej komponenty krustáceoplanktónu - veslonôžok - v stredovej zóne boli zastúpené iba pravé planktonty, najmä Thermocyclops oithonoides a Th. crassus (v Istragovskom ramene) spolu s niektorými ďalšími druhmi. Ramená typu plesiopotamál Z monitorovaných ramien, nielen v hlbšom ramene na Kráľovskej lúke pri Trstenej, ale aj v plytkom zvyšku ramena na Spornej sihoti pri Kľúčovci v krustáceoplanktóne dominovali typické planktonty, ba v prvom roku boli jeho jedinou zložkou (Vranovský, Illyová 1999). V ramene na Kráľovskej lúke to bola z perloočiek Diaphanosoma brachyurum alebo Bosmina longirostris, z veslonôžok Thermocyclops oithonoides, v ramene na Spornej sihoti medzi najčastejšie a aj v ročných priemeroch dominujúce druhy patrila perloočka Bosmina longirostris a veslonôžky Cyclops vicinus, Thermocyclops oithonoides, Th. crassus, Eudiaptomus gracilis., Eurytemora velox a tiež Acanthocyclops robustus. Po zátopách inundačného územia sa v tomto type ramien vytváralo spoločenstvo s dominanciou euplanktonických druhov, najmä veslonôžok Cyclops vicinus a Thermocyclops crassus. V iných obdobiach tu nadobúdali kvantitatívnu prevahu druhy charakteristické pre litorál, resp. plytké občasné (temporárne) stojaté vody: z perloočiek najmä Chydorus sphaericus a Ceriodaphnia reticulata, z veslonôžok Megacyclops viridis, Metacyclops gracilis, Eudiaptomus transylvanicus a Cryptocyclops bicolor. Pôvodné spoločenstvá zoobentosu Hlavný tok Dunaja Spoločenstvá nálevníkov (Matis, Tirjaková, 1995 a, b) sa ukázali byť pomerne chudobné z hľadiska druhového zloženia i abundancie. Zastúpené boli hlavne euryékne druhy (bakteriovorné – Cyclidium glaucoma, Aspidisca cicada, A. lynceus, Glaucoma scintillans a i.). V relatívne malom počte boli zastúpené planktonické druhy. Ostatné sledované zložky mikrozoobentosu a meiozoobentosu boli nachádzané sporadicky. Bolo to spôsobené nedostatkom vhodných substrátov, veľkou rýchlosťou prúdu, zanášaním kalom, vyplavovaním turbulenciami a nestabilitou výšky hladiny. Po záplavách Dunaja sa opakovane zisťovali inaktivácie jedincov (pravdepodobne vplyvom splavených toxických látok).V tomto období sa objavili na jednotlivých lokalitách aj zriedkavejšie sa vyskytujúce druhy napr. Ophryoglena flava, Tintinnopsis cilindrata, Stegochilum fusiforme, Frontonia anbigua, Strombidium turbo. Permanentná fauna makrozoobentosu mala v sledovanom úseku rieky v litorálnej zóne kvalitatívne jednotný charakter (Krno et al., 1999). Medzi dominatné druhy patrili - Eunapius fragilis (Porifera), Dendrocoelum lacteum (Turbellaria), Dina punctata (Hirudinea), Ancylus fluviatilis, Lymnaea ovata, Bithynia tentaculata (Gastropoda), Dreissena polymorpha, Sphaerium corneum (Bivalvia) (Košel, 1995a). Z máloštetinavcov to boli druhy z čeľ. Naididae a Stylodrilus heringianus (Lumbriculidae), zriedkavo sa vyskytovali druhy z čeľ. Tubificidae. Významný bol aj výskyt druhu Hypania invalida (Polychaeta) a Dikerogammarus haemobaphes a Corophium curvispinum (Amphipoda). Isté rozdiely vo faune boli na profile Kľúčovec, ktorý leží už poniže spádového zlomu Dunaja (Šporka, Krno 1995). V taxocenózach temporárnej fauny v litoráli Dunaja dominovali Baetis fuscatus, Heptagenia sulphurea, Caenis pseudorivulorum (Ephemeroptera), Hydropsyche contubernalis, H. bulgaroromanum, Psychomyia pusilla Brachycentrus subnubilus, Ceraclea dissimilis (Trichoptera). Oproti výskumom z 80. rokov (Krno, 1990) sme už nezaznamenali viaceré druhy podeniek - Heptagenia coerulans a rod Ecdyonurus a ďalšie druhy sa vyskytovali veľmi zriedkavo - Baetis vardarensis, Heptagenia flava, Ephemerella ignita a Potamanthus luteus. Kvantitatívne bola dunajská fauna najbohatšia pod vyústením ramien pri Gabčíkove, pričom bol výrazne bohatšie osídlený skalnatý substrát (Krno, 1995). Na skalnatom substráte dominoval druh H. bulgaroromanum, na štrkovitom H. contubernalis. Celkove prevládali filtrátory (Hydropsychidae, Brachycentrus). Mediálna časť hlavného toku Dunaja (r. km 1816), ktorú sledovala Ertlová (1968) bola osídlená hlavne máloštetinavcami (Nais elinguis, Chaetogaster crystallinus, Propappus volki, Rhynchelmis limosella a Stylodrilus heringianus), pijavicami (Erpobdella octoculata), pakomárovitými (Polypedilum sk. laetum, P. sk. scaleanum, Ablabesmyia sk. lentiginosa a Euorthocladius rivicola). Ramená typu parapotamál Na našom území sa z hľadiska štúdia mikrozoobentosu venovala pozornosť predovšetkým ramenám (Matis, Tirjaková, 1992; Tirjaková, 1992; Szentivány, Tirjaková, 1994). Z hľadiska štruktúry sledovaných skupín nie je možné sledované ramená hodnotiť ako celok. Spoločenstvá každého z nich vzhľadom na svoj charakter a meniace sa podmienky sa vyvíjali špecificky. Spoločným znakom všetkých sledovaných ramien pred napustením vodného diela je dlhodobý postupný pokles hladín vody, ich premena na ramená s prúdiacou vodou len počas vyšších prietokov v Dunaji. V období po ich odrezaní od hlavného toku sa tu vyskytovali bohaté spoločenstvá typické pre stojaté vody. Ramená so stojatou vodou a preplachované počas povodní sa vyznačovali širokým druhovým spektrom a abundanciou všetkých skupín mikrozoobentosu (Ciliophora, Mastigophora, Heliozoea, Amoebina). V permanentnej faune slepých ramien, sa pôvodne vyskytovali jednak rovnaké druhy ako v hlavnom toku Corophium curvispinum, Dikerogammarus haemobaphes (Amphipoda), Hypania invalida (Polychaeta), Stylodrilus heringianus a rody Psammoryctides a Potamothrix (Oligochaeta) (Košel, 1995a; Krno et al., 1999). V prietočnom ramene pri Istragove dominovali, ako zistila v roku 1966 Ertlová (1970), Oligochaeta (Potamothrix moldaviensis, Tubifex tubifex, Tubifex ignotus a rod Limnodrilus) a Chironomidae (Prodiamesa olivacea, Chironomus sk. thummi, Cryptochironomus sk. defectus). Vysokú abundanciu v hlavných ramenách Bačianskej ramennej sústavy v rokoch 1976 – 1978 na štrkopiesčitom dne mediálu dosahovala Dreissena polymorpha (Bivalvia), ktorej agregácie boli vyplnené jemnými sedimentami a osídlené osobitým bentickým spoločenstvom, druhové zloženie ako aj početnosť bentickej fauny bola vysoká (Šporka, Nagy 1998) Vysoké stavy síce spôsobovali dočasné ochudobnenie fauny, po krátkom období, cca 35 dní, sa pôvodné spoločenstvo dokázalo zotaviť. V rámci temporárnej fauny v prietočných ramenách Lichardová (1958), pred rokom 1960 zistila viaceré taxocenózy podeniek v ktorých sa pravidelne vyskytovali druhy – Potamathus luteus, Heptagenia sulphurea, Ecdyonurus aurantiacus, Baetis rhodani, B. fuscatus, Serratella ignita. Svedčí to o výrazne väčšej reofilnosti týchto ramien. Podobná situácia v dunajských ramenách bola s taxocenózou potočníkov (Mayer, 1935) – Rhyacophila pascoei, Agapetus sp., Hydroptila sp., Plectrocnemia sp., Neureclepsis bimaculata, Polycentropus flavomaculatus, Hydropsyche spp., Cheumatopsyche lepida, Setodes interruptus, Potamophylax latipennis, Halesus spp., Goera pilosa, Silo pallipes a Brachycentrus subnubilus. V ramenách Bačianskej ramennej sústavy v rokoch 1976 – 1978 bolo z podeniek zistených 6 taxónov a z potočníkov zástupcovia dvoch čeľadí, okrem toho z pakomárovitých bolo zistených 22 taxónov (Šporka, Nagy 1998). V rokoch 1991 – 1992 temporárna fauna prietočných ramien bola pomerne chudobná, zastúpená druhmi Cloeon dipterum, Caenis horaria, C. luctuosa (Ephemeroptera) a rodmi Ecnomus, Cyrnus, Anabolia, Athripsodes (Trichoptera). Z vážok (Majzlan, 1992) dominovali v parapotamáli druhy Calopteryx splendens a Lestes viridis. Spoločenstvo pakomárovitých sa vyznačovalo prítomnosťou druhov uprednostňujúcich tečúce typy vôd (Cricotopus bicinctus, Tanypus kraatzi) ako aj druhmi preferujúcimi pomalšie tečúce, alebo stojaté vody (Dicrotentipes spp., Polypedilum spp.) (Krno a kol., 1999). Ramená typu plesiopotamál V mŕtvych ramenách Dunaja sa vyskytovali pomerne stabilné spoločenstvá mikrozoobentosu (najmä na lokalitách Kráľovská lúka a Sporná sihoť). Na lokalite Kráľovská lúka vplyvom zníženia hladiny však dochádzalo v letných mesiacoch k prehrievaniu vody, čo malo za následok zmeny v štruktúre spoločenstva hlavne nálevníkov (masové premnoženie niektorých druhov, napríklad Coleps nolandi) (Matis, Tirjaková, 1995a,b) . V litoráli ramien typu plesiopotamál sa vyskytovalo jednak štrko-piesčité dno a jednak bahnité dno. Na štrko-piesčitom dne bolo druhové zloženie permanentnej fauny pestrejšie, vyskytovali sa tu okrem druhov z čeľ. Tubificidae aj druhy z čeľ. Naididae, teda druhy živiace sa nárastami, kdežto v bahnitom dne prevládali druhy z čeľ. Tubificidae, prípadne amfibiotické druhy čeľ. Enchytraeidae, veľké druhy Criodrilus lacuum a Eiseniella tetraedra (Nagy, Šporka, 1990). Druhove chudobné boli aj ostatné skupiny, pričom prevažovali ulitníky so 14 druhmi. Abundancia väčšiny druhov bola pritom veľmi nízka, jedine kôrovec Asellus aquaticus sa v roku 1992 zistil vo zvýšenom množstve (Šporka, Krno, 1995). Z temporárnej fauny sa tu vyskytovali hlavne stagnikolné druhy vážok Sympetrum flaveolum, Lestes barbarus, Cordulia aenea ale bol tu zistený semireofilný druh Platycnemis pennipes (Majzlan, 1992). Spoločenstvo pakomárovitých bolo druhove chudobné, pričom dominantné postavenie zaujímali pelofilné druhy Cryptochironomus defectus a Polypedilum nubeculosum (Krno et al., 1999). V ramene Žofín v roku 1971 medzi dominantné skupiny patrili Oligochaeta a Chironomidae Ertlová (1973). K rovnakým záverom dospeli aj Nagy a Šporka (1990) v rokoch 1981 – 1987 na ramene pri Kráľovskej lúke, kde najvyšší počet taxónov u permanentnej fauny bol zistený v litoráli (bez zárastov a so zárastami), mediálna zóna bola osídlená len chudobne. Mediálnu zónu s bahnitými sedimentami osídľovali v prevažnej miere larvy pakomárovitých, ktoré sa druhovo početne vyskytovali aj v litorálnej zóne ramena. Pôvodné ichtyocenózy v hlavnom toku a v ramenných sústavách Pôvodné ichtyocenózy v hlavnom toku a ramenných sústavách od ústia rieky Moravy po ústie rieky Ipľa opisuje Balon (1966), na základe prác Laboratória rybárstva v rokoch 1953–1961. Udáva existenciu 56 druhov rýb s komentárom ich výskytu. Výskumy v nasledujúcich troch desaťročiach však ukázali, že počet žijúcich druhov v tomto úseku Dunaja je výrazne vyšší, takže je v porovnaní s ostatnými tokmi na Slovensku druhovo najbohatším. Najnovší zoznam druhov rýb slovenského úseku Dunaja obsahuje 76 položiek. Z toho 61 druhov je pôvodných, 10 (11) introdukovaných exotických a 3(4) druhy sem invadovali z dolných úsekov Dunaja (Holčík 2003). V porovnaní so všetkými slovenskými riekami ichtyocenóza tohoto úseku Dunaja je druhovo najbohatšia. Autor si to vysvetľuje dvomi faktormi: 1) skutočnosťou, že ide o začiatok podhorskej zóny, presnejšie, o prechod medzi podhorskou a nížinnou zónou – medzi hyporitrálom-epipotamálom a metapotamálom v zmysle klasifikácie Illies a Botosaneanu (1963); 2) zmenou spádu koryta Dunaja, ktorá zapríčinila vznik rozsiahlej vnútrozemskej delty s komplikovaným systémom ramien. Toto spôsobuje, že na relatívne krátkom úseku Dunaja sa nachádzajú veľmi rôznorodé typy prostredia, čo vytvára druhovo bohaté spoločenstvo rýb. Antropické zásahy ovplyvnili ichtyocenózy v Dunaji. Výstavba vodného diela „Železné vráta“ na dolnom Dunaji zamedzila výskyt migrujúcich druhov. Naviac v Dunaji pribudli introdukované – allochtónne druhy. V období rokov 1970 – 1980 pri kulminácii organických a toxických polutantov došlo skoro k vymiznutiu lososovitých druhov citlivých na znečistenie, ako aj Cottus gobio, Phoxinus phoxinus, Alburnoides bipunctatus a Barbus barbus. Zlepšenie kvality vody v rokoch 1980 -1990 sa prejavilo k ich opätovným výskytom. Od r.1994 sa začali objavovať aj nové druhy rýb z čeľade Gobiidae, predtým známe iba z dolných úsekov Dunaja. Konektivita hlavného toku Dunaja s ramennými systémami priaznivo ovplyvňovala ich druhovú diverzitu. Ako konštatuje Holčík (2001), hodnoty počtu druhov, druhovej diverzity a vyrovnanosti sú v hlavnom toku vyššie a laterálne smerom od hlavného toku ku okraju vnútrozemskej delty vykazujú pokles. Príčinou sú rozdielne podmienky prostredia v ekosystémoch jednotlivých typov vôd vnútrozemskej delty. Nebyť vnútrozemskej delty, druhová diverzita v hlavnom toku by bola nižšia. Spôsobuje to hlavne existencia vhodných neresísk v ramenách a vhodné refúgia hlavne počas povodní, ako aj dobrá potravinová ponuka ramien. Vyhodnotenie pôvodného stavu ichtyocenóz na monitorovacích plochách a v Dunaji, tesne pred prehradením Dunaja v roku 1992, podáva Černý (1999).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}