V.2. BiotaV.2.1. Danube and the Danube surrounding biota in the course of timeCompiled by Iľja Krno and Mikuláš J. Lisický The construction and operation of the Gabčíkovo project represents a significant milestone in the functioning of the Danube ecosystem and its environment. An objective evaluation of the existing and potential future changes is impossible without knowledge and analysis of the pertinent trends in the past centuries. The first attempt to evaluate the Danube and its natural environment was done by A. F. Marsili [10], while J. B. Grossinger brought the first concrete data on the Danube fauna already in the late 18th century. The first reliable data on the occurrence of concrete plant species come from the first half of 19th century (S. Endlicher for surrounding of Bratislava). The first reliable faunistic data come from the turn of 19th to 20th century. However, up to the mid-20th century, most of the publications had just a floristic or faunistic character and cannot be used for purposes of evaluation at the ecosystem level. Kalivodová, Janotková and Kis-Csájiová [4] compiled selective bibliography of zoological papers Already in the 1930s the Hydrobiological Station of the Šafarík Learned Society existed in Šamorín, but it was purely faunistically oriented. The first ecological studies in this stretch of the Danube (inclusively of the Austrian and Hungarian stretch) started to be sporadically made in the 1940s in the frame of geobotanical investigations (F. Hartmann, R. Soó, B. Zólyomi). More systematic works originate from the 1950s (J. Berta, A. Jurko, A. Mezera, E. Wendelberger), when also the post-war generation of Slovak hydrobiologists and ichthyologists (E. Balon, I. Bastl, J. Brtek, M. Ertl, E. Ertlová, J. Rothschein, M. Vranovský, J. Holčík) began to work in this area. At the same time, the first biological data on Danube pollution were collected (V. Mucha, L. Hanuška, R. Jacko) [14]. The first regional attempt to summarize known limnological data was done by R. Liepolt [7]. It is obvious that most of the relevant data, which could serve as a basis for evaluating this ecosystem, originated from recent decades, from the time, when the Danube was already considerably canalised and its connectivity with the side water bodies was significantly reduced. Thus, a comparison is possible, but only with the consciousness that we compare two anthropogeneously influenced states of natural environment in a period, which is not long enough for generalization. An image about secular changes can be done only roughly on the basis of paleobotanical (E. Kripel) and Quaternary-malacozoological works (V. Ložek). A map of potential natural vegetation was elaborated for plant communities [11], which can be used for an evaluation of the degree of naturalness of biocoenoses, although without detailed ecological analyses. The molluscs also may give information about the natural mollusc fauna of an area based on regio-types [8]. Recently, initiative papers combining data from historical geomorphology, paleobotanics and malacozoology (see for example P. Pišút and T. Čejka, in this issue) arose in the Slovak part of the Danube area. The first ecosystem studies directly related to the Danube hydraulic structures were done in the 1980s in Austria [1,13]. Almost one year before putting the Gabčíkovo project into operation, a group of experts under the leadership of J. Holčík summarized the existing knowledge about the broader Danube area and formulated certain prognoses related to the project [3]. The 1990s was decisive for data collection and the creation of a complex image about the Danube ecosystem and its floodplain. The observation system to monitor biota influenced by the Gabčíkovo hydraulic structures is globally unique, not only because of the number of collaborating specialists (55 in 1990), but also because of the unprecedented concentration on one problem at the same time. The system was built up on the principle of common monitoring plots and their representativeness for the existing natural biocoenoses and expected environmental changes potentially caused by the hydraulic structures. Despite the later reduction of the monitored groups just at the time changes began appearing (1993-1995), the initial state is relatively well documented. At present, we can evaluate both 15- and 10-year periods. In the following text we shall focus especially on trends, while for the details we will refer to the monograph [12] and the rich bibliography given in this monograph. Biota of the Danube and its side water bodies, as well as the adjacent floodplain, has already been under strong anthropogeneous pressure for more then one century. Cutting off the arms of the main stream, sinking the river bottom because of dredging for navigation purposes, flood protection and gravel exploitation, the reduced bedload (transport of sediments) from Austria, and some other events, had substantially changed the conditions for development of hydrocoenoses in the Danube floodplain even before putting the Gabčíkovo project in operation. A result of this has been, for example, the extinction of more than 90% of the species of Plecoptera, more than 50% of the mayflies and more than 30% of the caddisflies [6]. When compared with the more remote past, canalisation of the Danube, concentration of water into one main canal, and flood-protection measures, as well as the construction of various dams and hydraulic structures in the upstream Danube stretch, had already substantially changed the hydrological dynamics of the Danube, floodplain, and its alluvial fan. Two natural seasonal flooding of the inundation area, in spring and summer, already do not occur in Slovak territory ([9], page 30). This resulted in a considerable reduction in the amount of sediments and, consequently, in an accelerated terestrialization of the arms. Many birds have ceased to breed there, namely: purple heron (Ardea purpurea), Grey Heron (Ardea cinere), Great White Egret (Egretta alba), Great bittern (Botaurus stellaris) and Garganey (Anas querquedula), (Bohuš, in this issue). Changes caused by the Gabčíkovo project Changes caused by the Gabčíkovo hydraulic structures, as well as changes without any connection with the operation of the hydraulic structures, were evaluated in detail in various publications, mainly [9, 12]. They brought information about the initial composition of aquatic biota and changes in its composition after more years of monitoring. The concrete data supporting our conclusions are in those papers. Aquatic communities in general After putting the Gabčíkovo hydraulic structures into operation, important changes started in the hydrological regime both in the former main channel, and in the arm system. This was especially manifested by changes of the macrozoobenthos communities [5]. The evaluation of changes was based on model groups. The benthic fauna and zooplankton are used as indicators of immediate and medium-term changes in processes running in the aquatic ecosystem. Fish and macrovegetation are used rather for evaluation of the medium- to long-term changes. After finishing the project, the absence of annual floods reduced the number of habitats serving as spawning places for fish, which caused changes in the structures of fish communities. It also explains the drop in their productivity (J. Černý, in this issue). The importance of natural flooding on fish fauna in the Old Danube, and especially in its arms, can be documented by the increase in recorded cumulative abundance and species diversity of fish after the flood in August 2002. Similar experiences are also described by E. Bulánková, I. Krno (in this issue). If conditions for hydrofauna in the within-dike zone would not be changed, and aquatic animals could not migrate between the eupotamal and the arms or between individual arms (flooding of Istragov, artificial spawning places), their natural reproduction would not be possible. In such a case, in spite of systematic fish stocking, we expect a further decline of fish production down- and upstream of the Gabčíkovo hydraulic structures, as well as the decline of fish catches in this stretch of the Danube. The other important impact results from the fact that the hydroscheme reduced the overall surface of the floodplain (within-dike zone in Čunovo reservoir), destroyed previous spawning places, and inhibited the migration of fish and macrozoobenthos (J. Černý, see more in this issue). Therefore we propose specific measures to improve the present state and to more effectively monitor fish. Some fish species, which were common in the Danube in the past, have been disappearing in the last decades (Cottus gobio, Gobio albipinnatus, Barbatula barbatulus), some of them are extinct (Gobio uranoscopus, Gobio kessleri). Values of all hydrobiological metrics show that the natural and simulated floods favorably influence the development of hydrocoenoses, and represent a necessary condition to preserve a good ecological state of the aquatic ecosystems in the area of the Gabčíkovo project (E. Bulánková, I. Krno, in this issue). The former Danube main channel (old riverbed) In the Old Danube the decreased discharge resulted in a decline of stream velocity and in the stabilization of gravel sediments, during usual discharges. The shallow ripal has widened and shifted towards the mid of the channel. Due to this many natural covers for aquatic animals have disappeared. The cumulative abundance and species diversity of hydrofauna decreased. Ripal of the old riverbed has started to be gradually colonized especially by juvenile individuals of fish (J. Černý, in this issue). The shallow water and increased transparency (sedimentation of suspended solids in the Čunovo reservoir) has enabled the development of benthic algae, which is an increased food resource for scrapers, especially for Copepoda, Cladocera, Oligochaeta and caddisflies (F. Šporka; I. Krno, in this issue). Later, after 1999, the bottom started to be covered by a thick layer of silty-clayish sediments. This resulted in a decrease of scrapers abundance (V. Košel; E. Bulánková, I. Krno, in this issue) because of the decline of algal growths as their main food. The mayflies (Ephemeroptera) almost disappeared from the Danube benthic fauna. Among the caddisflies (Trichoptera), collector-filterers of the family Hydropsychidae predominated in the Danube before 1999, but at present the collector-filterers of the genus Brachycentrus predominate. Changes in hydrological and trophical conditions in the Old Danube were manifested by the increased abundance of some original and typical lenitic species of Oligochaeta and dragonflies, which earlier occurred mainly in the arms. The largest proportion of reophilous and semireophilous taxa was recorded in the Old Danube riverbed in 1999 after a series of simulated floods in 1995, 1997 and 1998. Later, the proportion of semireophilous taxa increased, indicating a decline in stream velocity in the Old Danube (E. Bulánková, I. Krno, in this issue). After the water diversion, part of the euplanktonic crustaceans decreased in the Old Danube. This is a result of the decline of zooplankton reproduction in the arm system and its limited drifting into the old riverbed. However, in the last five years, the true plankton reappeared in the old riverbed. Now they are not represented by the cladocerans drifting in from the arms as in the past, but their occurrence is probably autochtonous. It is still to be verified to what degree the old riverbed is enriched by cladocerans that drifted in from the Čunovo reservoir. M. Illyová (in this issue), states that the character of the species composition of Cladocerans in the old riverbed (eupotamal) approximates the parapotamal, despite the fact that the typical feature of the parapotamal, the stagnant period, does not exist here. The slowing down and reduced flow velocity in the Old Danube and the creation of more favourable conditions for development of the planktonic community can explain the occurrence of these species. As this stretch is flowing water, the occurrence of these cladocerans is sporadic and no richer planktonic communities were observed. Upstream of the mouth of the tailrace canal, in the area of Istragov, a backwater stretch has arisen and the stream velocity was further reduced. Sedimentation of silt particles increased, especially at the time of increased discharges in the Old Danube. The originally gravely bottom was covered by a layer of clayish-sandy sediment, which caused a change of the original benthic community. In the profile at Istragov, the Amphipods and aquatic insects (excepting the Chironomids), being earlier dominant, have disappeared. Yet new representatives of Oligochatea, especially of the genus Limnodrilus, have appeared here (Šporka, in this issue). A significant change in mollusc, fauna accompanied by an outbreak of Theodoxus fluviatilis, was also recorded. The future existence of the original Danube fauna of aquatic molluscs is problematic (Košel in this issue). As to the development of saprobity (Nagy in this issue) in the Danube at Bratislava and Medveďov in 2001-2005, in the Čunovo reservoir left side at Kalinkovo, and in the arms system, a balanced degree of beta-mezosaprobity was recorded. In the Danube at Dunakiliti and Sap, a trend of moderate improvement from alpha-mezosaprobity to beta-mezosaprobity was observed; probably due to this, there has appeared for the first time the rare and endangered species of Plecoptera - Xantoperla apicalis a Taeniopteryx schoenemundi (E. Bulánková, I. Krno, in this issue). Similarly, the permanent presence of alpine bullhead (Cottus gobio), which requires a high content of oxygen and clear water, documented a very good water quality after the Danube damming (Černý in this issue). In the Čunovo reservoir, in the part close to Šamorín, a trend of moderate worsening from beta-mezosaprobity to alpha-mezosaprobity was recorded. The main communities indicating beta-mezosaprobity in the profiles monitored are lithoreophilous and psammophilous zoocoenoses; on the contrary the psammophilous and pelophilous benthic zoocoenoses indicate alpha-mezosaprobity. On the shallow marginal parts of the Old Danube, which are not permanently flooded, the bottom is covered by a natural succession of softwood floodplain forests. This represents an interesting research object, as the natural spontaneous reproduction of such stands was almost impossible in the last decades. On the other hand, such growth increases hydraulic roughness of the riverbed and the risk during flood discharges. Arm system The gradual adjustment of ground water levels and the following adjustments of water levels in different types of shallow water bodies resulted in a change of its character. Alternation of temporal denudation and flooding of the bottom ended here, and development is now heading to a division into permanently flooded or permanently dry formations. These factors gradually reduce the natural function of the inundation (J. Černý, in this issue). Supplying the arm system by permanently discharging water through step-like arranged cascades with backwater creates a different character. In the initial stage, i.e. 10 years ago, the most characteristic feature of fauna of the permanently discharging arms was its approximation to the benthic fauna of the former old riverbed, hence colonization by reophilous species. At present, the relatively long lasting isolation of arms from the main stream results in an increasing representation of littoral zooplankton (M. Illyová, in this issue) bound to macrophytes, whose mass spreading started in 2005, after extensive clear-cuttings in the vicinity of the arms (Bodícke rameno, Kráľovská lúka). The former, temporarily through flowing arms of the plesiopotamal types are not washed by water to the degree necessary to inhibit their being overgrown by macrovegetation and terrestrialization. Therefore the zoobenthos species trophically bound on macrophytes predominates in these arms (F. Šporka; E. Bulánková, I. Krno; M Illyová, in this issue). Plants die off in autumn. Large amounts of biomass, and it decay in winter, especially under ice, is negatively influencing the wintering hydrofauna. Thus, besides oxygen shortage caused by overheating in summer in stagnant waters, a winter oxygen shortage also occurs and reduces the species diversity of hydrofauna. A particular problem is the area of the Istragov Island, which became dry because not supplied with water from the bypass canal. Although a system of intake devices already exists, it has never worked. Changes also occurred in the structure of local fish communities. The eurytopic, especially phytophilous and, in the within-dike zone, commonly occurring species with a wide ecological valence started to predominate in monitoring plots. The relative abundance of reophilous fish species, which earlier formed the major part of fish communities in the arms even in periods out of communication with the former main channel, has now become subdominant and receding (J. Černý, in this issue). The increase of species diversity in planktonic crustaceans in the river arms is also a consequence of the natural process of overgrowing and terrestrialization of these water bodies. This process is also accelerated by the frequency and intensity of simulated or natural floods, which are much less that the number of natural floods in the past. This inevitably leads to extinction of these habitats and to the disappearance of rich fauna of planktonic crustaceans. The discharging and dynamic regime of the arms has turned into a static regime, with the individual parts overgrown by macrovegetation. Therefore, at least a temporal communication of these water bodies with the former main stream of the Danube should be provided. This is important especially at periods of flood discharges, to wash sediments, to enrich them by nutrients, and to slow down the process of their being overgrown, shallowed and terrestrialized. Dynamics of surface water regime and lateral erosion A significant change in the ecosystem was a decrease of water level fluctuation dynamics in the shallow side water bodies, which almost regularly provided periods of local occurrence of shallow water in the arms (most important in late summer and autumn). As they represented significant feeding areas for more waders (for example Grey Heron, Great White Egret, Black Stork), the absence of shallow water in the arms has reduced the availability of food for this trophical guild of birds, just in the critical period between the fledging of chicks and migration. Sometimes it has led to the loss of wetland habitats character suitable for waders (M. Bohuš, in this issue). The fact that the highly endangered stenoecious Spoted Crake (Porzana porzana) was not recorded in the vegetation seasons of the last few years can be connected with the gradual extinguisment of wetland habitats diversity. The most humid types of natural floodplain forests (Salici-Populetum myosotidetosum and Salici-Populetum phragmito-caricetosum) have become drier, which has caused adaptive changes in the zoocoenoses living there (T. Čejka, in this issue) as represented by the molluscs. M. Bohuš (in this issue) found out that an increase of coverage of herbal stratum secondarily caused a linear increase of abundance of Blackcap (Sylvia atricapilla) and River Warbler (Locustella fluviatilis), with a simultaneous decline of abundance of Redstart (Phoenicurus phoenicurus). Extinguishing lateral erosion reduces breeding opportunities of Kingfisher (Alcedo atthis) (M. Bohuš, in this issue) and Sand Martin (Riparia riparia). Inundation area Changes in terrestrial communities were already manifested in the first year after the Danube diversion, in a different intensity in dependence on mitigating measures and the decline of ground water level. In the first half year the arm system was first emptied and later permanently supplied with water from the intake structure at Dobrohošť. In the inundation area on the Slovak side, three parts remained out of reach of supplying water. In each of them, changes could be observed with some specific features: - Area upstream from the intake structure between Hrušov and Dobrohošť (Dunajské kriviny), where a drop of ground water level into the gravelly substrate appeared immediately after the Danube diversion. The original softwood floodplain forest disintegrated. After a temporary occupation of the opened dry places by a herbal stratum, resulting in changes in the epigeic fauna (Z. Šustek, in this issue), the area was overgrown by tolerant shrubs. Because this site represents one of the monitoring plots, all changes are well documented in databases and photographs [9, 12]. - The riverbank strip along the Old Danube (several tens of meters, locally more than 100 m), due to the drain effect of the Old Danube riverbed. The gradient is growing downstream according to the difference between the water level in the branches and in the Old Danube. In the arm system weirs inhibit the decline of water level. In the old riverbed the discharge corresponds to a quarter of the original one in vegetation season. In regard to the fact that prognoses of changes in these sites were unambiguous, in two clear-cut plots trees typical of the hardwood floodplain forests (oak, ash) were experimentally planted. - The area between the tailrace canal and the old riverbed (Istragov – Riečiny). In this area, the unfavourable effect on terrestrial communities is partly buffered by backwater in the old riverbed, upstream of the confluence with the tailrace canal. In the areas where the soil moisture is influenced by the arm system supplied with water, and where simulated floods are possible, the changes recorded have an adaptive character. The dynamics of microclimate fluctuations are inhibited here and altitudinal differences in terrain shape start to play a more significant role. Especially the terrain depressions represent islets of increased humidity. The species richness of most forest phytocoenoses was impoverished by 4-6 species (Uherčíková in [9]. In the first years after the Danube diversion (1993-1994), the ground beetle communities were characterized by a strong increase in abundance of two or three less hygrophilous species (Pterostichus melanarius, Pterostichus niger, locally also Carabus granulatus). Simultaneously, the abundance and biomass of all the more hygrophilous species of Carabidae a Staphylinidae started to decline. The most affected species were Carabids Platynus assimilis, Patrobus atrorufus, Oxypselaphus obscurus, Europhilus fuliginosus, Europhilus micans and Staphylinids Oxytelus rugosus and Tachinus rufipes. Despite a decline of abundance and biomass of more hygrophilous species, the cumulative abundance and biomass of communities of both families increased. In the next period (1995-1996), quantitative representation of the most hygrophilous species continued to decrease, but representation of the three more tolerant Carabid species also declined. The communities started to be invaded by moderately hygrophilous Epaphius secalis and the xenocenous field and ruderal species Trechus quadristriatus. Size of the total catche of individuals of both families dropped from about 3000 to 1000. In 1997, the microclimatical conditions moderately improved, which allowed the more hygrophilous component of the community of Carabidae and Staphylinidae to stregthen its position (Z. Šustek, in this issue). Surroundings of the Čunovo reservoir The increased level of ground waters in the surroundings of the Čunovo reservoir was favourably manifested by the occurrence of aquatic macrophytes in sites, which are not intensively influenced by water streaming. Some of the earlier vanishing aquatic habitats were again supplied with water (S. Kubalová, in this issue). In the within-dike zone, the vegetation of aquatic habitats is influenced by hydrological conditions in individual sites, which result from the local geological and geomorphological conditions, as well as from climatic conditions in the given year. In those sites, which are flooded during major part of the vegetation season (especially in summer) by stagnant or slowly flowing water, the aquatic macrophytes find optimal conditions for their existence. Changes in macrophytic vegetation after the Danube damming are not significant and are subject to regular fluctuations resulting from changes in hydrological regime. However, these fluctuations are characteristic of the Danube floodplain. Changes supported by operation of the Gabčíkovo hydraulic structures Measures undertaken in the within-dike zone in favour of the ecosystem (supplying the arm system with water and the possibility to manipulate with water levels) also meant an easy accessibility of the major part of this zone, not only by walking, but also for vehicles and different mechanisms. Eventually, the forest economy intensifies and also affects those plots, which were minimally affected due to their bad accessibility. Old and hollow trees are intentionally removed, which results in the gradual elimination of hollow-nesting breeders. Easier accessibility of forests stands and intensification of forest economy also liquidate nesting opportunities of Black Stork (M. Bohuš in this issue). Also monitoring of molluscs shows that poplar monocultures have a significance for survival of autochtonous mollusk communities only in older age classes, in which two thirds of terrestrial mollusks (32 species) are able to exist (Čejka, in this issue). Neophytic plants with a tendency to acclimatize started to spread in the area (Uherčíková in [9]). This results from a worsening of moisture conditions, intensifying of forest economy, but also from increased pressure of recreation activities, which even include gardening. Allochtonous decorative plants, trees and shrubs (for example arbor vitae, pines) are intentionally planted and the vicinity of weekend houses is ruderalized. Nitrophilous vegetation also spreads (Uherčíková in [9]). Unsufficiently regulated recreation on excavation pits and their shores, and a larger frequancy of „tourists“ in floodplain forests represent a danger. In the past, the recreation cottages were constructed on pillars due to regular floods, but at present they are constructed directly on the ground surface and some of them are even provided with cellars! This produces pressure by the recreation loby to exclude completely the simulated floods in the within-dike zone. Changes in natural environment independent of the Gabčíkovo hydraulic structures Since 1996, four new allochtonous fish species of the Gobiidae family (Neogobius kessleri, N. melanostomus, N. fluviatilis, N. gymnotrachelus) were recorded in the Slovak stretch of the Danube. In connection with the expansion of these species, we recorded the extinction of populations of Barbatula barbatula and Cottus gobio, as well as the decline of Gobio albipinnatus. We presume that the spreading of these allochtonous species, can have other, currently unknown, impacts on fish fauna. A similar mass spreading of some invertebrate species was also observed by V. Košel and M. Illyová (in this issue). Among the aquatic plants, spreading of the allochtonous species Elodea nuttallii (introduced by aquarists from North America) has an invasive character (Kubalová in this issue). In the monitored area, the White-tailed Sea Eagle (Haliaeetus albicilla) reappeared as a breeding species and three new breeders - Mediterranean Gull (Larus melanocephalus), Yellow-legged Gull, (Larus cachinnans) and Mute Swan (Cygnus olor) - started to occur here. On the contrary, the population size of Redstart (Phoenicurus phoenicurus) considerable decreased in the whole Danube floodplain in Slovakia, or locally this species has not appeared at all. Strong changes in the population sizes of a further three bird species (Phalacrocorax carbo, Ph. pygmaeus and Milvus migrans) have a regional character (M. Bohuš, in this issue). Improved water quality in the Danube created suitable conditions for the restoration of populations of the endangered mollusc Esperiana esperi, and also supported the spreading of the allochtonous mollusk Theodoxus fluviatilis (V. Košel, in this issue). References
V.2.2. Monitoring of saprobity based on composition of macrozoobenthos in the Danube, Čunovo reservoir and the branch system between Bratislava and Medveďov in 2002-2005 Štefan Nagy In addition to other components of hydrocoenosis, monitoring of macrozoobenthos in the area of the Gabčíkovo hydraulic structures has been carried out in our laboratory since 1994. In 1994-1998 we focused mainly on the Čunovo reservoir and put emphasis on the production parameters, abundance and biomass of macrozoobenthos, i.e. on its qualitative and quantitative structure, but without any relation to saprobity. Since 1999, we enlarged the network of monitored profiles on the downstream end of the arms system at Gabčíkovo, the Danube riverbed at Sap and the hydropower station tailrace canal. In addition to production parameters we also dealt with the relationship of the macrozoobenthos structure to the saprobity and to the establishment of the saprobic index. In 2002, we enlarged the network of monitoring places, in particular to include profiles in the Danube (Bratislava, Dunakiliti, Dobrohošť and Medveďov) and at the beginning of the river arm system at Dobrohošť. The tailrace canal was abandoned. Monitoring the production parameters has been stopped, with key emphasis being put on monitoring the relation of the macrozoobenthos structure to the saprobity. Methods Samples for establishment of the macrozoobenthos saprobic index were taken according to methods presented in the norms STN EN 27828 (Methods of biological sampling. Instruction for sampling benthic macroinvertebrates by hand net), STN EN ISO 9391 (Sampling of macroinvertebrates from deep waters – Instruction for use of colonization, quantitative and qualitative samplers) and STN ISO 5667-6 (Water quality. Sampling part 6: Instruction for sampling in rivers and brooks). Based on identification of the recorded indicative species of macrozoobenthos, we calculated saprobic indices and established the saprobity of macrozoobenthos according to the still valid norm STN 83 0532 (Biological analysis of surface water; Establishment of benthos and the saprobic index according to Pantle and Buck). The samples were taken three times a year in the spring, summer and autumn, predominantly in April, July and October, depending on hydrological conditions. The possible shifts in sampling timing were caused foremost by high water levels, which made relevant sampling impossible. Monitoring profiles and sampling sites Monitoring sites are: - The Danube – Bratislava, two sampling sites, separately on the left and right riverbanks at the Nový most bridge (rkm 1868.7) in a lotic habitat. - The Danube – Dunakiliti, one sampling site on the left riverbank at the weir (rkm 1843.1), both in lotic and lenitic habitats. - The Danube – Sap, one sampling site on the left riverbank (rkm 1812.5), both in lotic and lenitic habitats. - The Danube – Medveďov, one sampling site at the top of the stony groin downstream of the road bridge (rkm 1806,0) in a lotic habitat. - The Čunovo reservoir – Kalinkovo, two sampling sites (rkm 1855), at the streamline and on the left riverbank in lotic habitats. - The Čunovo reservoir – Šamorín, two sampling sites (rkm 1847), separately in the right and left side bays in lenitic to moderately lotic habitats. - Arm system – Dobrohošť, at the beginning, downstream from the intake structure, in a lotic habitat. - Arm system – Gabčíkovo, at the downstream end of the Bačianske rameno arm at the “J” arm weir, upstream from the mouthing into the Danube, both in lotic and lenitic habitats. We present the results from profiles where we took samples from two places below, with average values from both sampling sites and values from each sampling site: L – left side; R – right side; K – stream line. Description of results In spring, the saprobic index ranged from 1.88 (Danube – Medveďov, in 2003) to 3.04 (Čunovo reservoir – Šamorín, in 2004). Values of the saprobic index indicated predominantly the level of beta-mezosaprobity. The alpha-mezosaprobity level was recorded in spring only in the Šamorín part of the Čunovo reservoir in 2002 (3.04; L-3.03; R-3.04), 2003 (2.53; L-2.49; R-2.56) and 2004 (2.65; L-2.41; R-2.88), as well as in the Danube at Dunakiliti in 2002 (2.69). When evaluating the changes of average saprobic indices from the four-year period along the longitudinal profile downwards in the Danube, it is possible to state that in the spring the saprobity: - in the Danube at Bratislava reached an average beta-mezosaprobity (2.33; L-2.21; K-2.44), - in the Kalinkovo part of the Čunovo reservoir, alpha-mezosaprobity (2.67; L-2.77; R-2.56), - in the Danube at Dunakiliti, a worse beta-mezosaprobity (2.27), - at the beginning and at the end of the arm system at Dobrohošť and Gabčíkovo coincidentally, an average beta-mezosaprobity (2.08), - in the Danube at Sap, a worse beta-mezosaprobity (2.11), and, at Medveďov, once more an average beta-saprobity (2.07). In summer, the saprobic index ranged from 1.81 (arm system at Dobrohošť in 2005) to 2.88 (Čunovo reservoir at Šamorín in 2004). Again, the values indicating the level of beta-mezosaprobity predominated. The level of alpha-mezosaprobity was recorded, similarly to in spring, in the Čunovo reservoir at Šamorín in 2003 (2.57; L-2.69; R-2.44), in 2004 (2.88; L3.13; R-2.63) and in 2005 (2.63; L-2.72; P-2.54) as well as in the Danube at Dunakiliti in 2002 (2.79). From the perspective of changes along the longitudinal profile in the monitored area, in summer we recorded the saprobic index: - in the Danube at Bratislava, average beta-mezosaprobity (2.03; L-2.00; R-2.05), - in the Čunovo reservoir at Kalinkovo, worse beta-mezosaprobity (2.28; L--2.21; R-2.34), - in the Čunovo reservoir at Šamorín, moderate alpha-mezosaprobity (2.55; L-2.66; R-2.43), - in the Danube at Dunakiliti, worse beta-mezosaprobity (2.36), - at the beginning and end of the arms system at Dobrobošť and Gabčíkovo almost coincidentally, average beta- mezosaprobity (2.01 and 1.95, respectively), - in the Danube at Sap, worse beta-mezosaprobity (2.21), and, at Medveďov, again an average beta-mezosaprobity (2.09). In autumn, the saprobic index values ranged from 1.98 (coincidentally at the beginning and end of the arm system in 2005 and 2002) to 2.66 (Čunovo reservoir at Šamorín in 2003). In autumn the level of beta-mezosaprobity also predominated in most profiles. The level of alpha-mezosaprobity was again recorded in the Čunovo reservoir at Šamorín in 2003 (2.66; L-2.74; R-2.57), 2004 (2.61; L-2.70; R-2.52) and 2005 (2.51; L-2.46; R-2.56), as well as in the Danube at Sap in 2002 (2.51). When evaluating changes in saprobity along the longitudinal profile, we recorded: - in the Danube at Bratislava, average beta-mezosaprobity (2.06; L2.06; P2.05), - in both parts of the čunovo reservoir at Kalinkovo and Šamorín, worse beta-mezosaprobity (2.22; L-2.20; R-2.24 and 2.47; L-2.51; R-2.43, respectively), - in the Danube at Dunakiliti, worse beta-mezosaprobity (2.12), - at the beginning and end of the arm system at Dobrohošť and Gabčíkovo coincidentally as in the summer, average beta-mezosaprobity (2.01 and 2.00 respectively), - in the Danube at Sap, worse beta-mezosaprobity (2.29), and at Medveďov an average beta-mezosaprobity (2.07). As for the annual average, the values of saprobic index fluctuated in the range of 1.95 (beginning of the arm system at Dobrohošť in 2005) to 2.71; L-2.90; R-2.52 (Čunovo reservoir at Šamorín in 2004). According to annual averages, we recorded the following values along the longitudinal profile: - in the Danube in Bratislava, average beta-mezosaprobity (2.05; L-2.05; R-2.04), - in the Čunovo reservoir at Kalinkovo, worse beta-mezosaprobity (2.28; L-2.21; R-2.34), - in the Čunovo reservoir at Šamorín, moderate-alpha-mezosaprobity (2.56; L-2.65; R-2.47), - in the Danube at Dunakiliti, again worse beta-mezosaprobity (2,25), - at the beginning and end of the arm system at Dobrohošť and Gabčíkovo, almost identical average beta-mezosaprobity (2.03 and 2.02 respectively), - in the Danube at Sap, again worse beta-mezosaprobity (2.20), and, at Medveďov, worse beta-mezosaprobity 2.08. The course of recorded average values of saprobic index in individual years is given in Fig. 1, Fig. 2, Fig. 3, Fig. 4 and Fig. 5. From the viewpoint of development of saprobity in individual profiles during the 4-year period it can be stated that a balanced degree of average beta-mezosaprobity occurred at the profiles in the Danube in Bratislava, at Medveďov, and at the beginning and end of the river arms system. A balanced degree of worse beta-mezosaprobity was in the Čunovo reservoir at Kalinkovo, and in the Danube at Dunakiliti. At Sap we recorded a trend of moderate improvement from alpha-mezosaprobity to worse beta-mezosaprobity, whereas there was a trend of moderate worsening from worse beta-mezosaprobity to alpha-mezosaprobity in the Čunovo reservoir at Šamorín. The recorded values of saprobic index and the degrees of saprobity result from the qualitative and quantitative composition of macrozoobenthos, which is influenced especially by hydrological regime. This dominant complex of abiotic factors determines the formation of bottom habitats inhabited by specific macrobenthic zoocoenoses with dominant indication species. In profiles with relatively strong water current, which are represented by the profile in the Danube at Bratislava, Medveďov, and at beginning of the arm system at Dobrohošť, a sandy-muddy bottom arises, which is inhabited by psammopelophilous zoocoenosis. The dominant species in this community are Hypania invalida, Lumbricidae g.sp., Nais sp., Lithoglyphus naticoides, Bithynia tentaculata, Sphaerium rivicola, Sialis lutaria, Procladius choreus and Chironomus spp., which indicate worse beta-mezosaprobity to polysaprobity. The rocky-gravely bottom in a strongly lotic environment downstream from the weirs with well-oxygenated water is inhabited by lithoscorioreophilous zoocoenoses, indicating a better to average beta-mezosaprobity. Thus, the resulting values for these profiles correspond to average saprobity at the end of the arm system at Gabčíkovo, with worse beta-mezosaprobity in the Danube at Dunakiliti and Sap. The specific hydrological conditions of the Čunovo reservoir, characterized by a sudden widening of the riverbed with accompanying slowing down of the stream, created the conditions for the occurrence of different habitats in the transversal profile. At Kalinkovo two sampling sites were monitored, representing two different habitats. The first habitat is a shallow littoral of a left-side bay with slower water stream, with rough gravely bottom and frequent surge. It is inhabited by skorioreophilous zoocoenosis, dominated by Dikerogammarus villosus, Corophium curvispinum, Cricotopus spp. a Phytochironomus fodiens and indicating better to average beta-mezosaprobity. The second habitat is represented by a strongly streaming medial with medium depth (3-6 m), a gravely-sandy bottom. It is inhabited by a skoriopsammophilous zoocoenosis indicating worse beta-mezosaprobity to better alpha-mezosaprobity. The dominant species of this zoocoenosis are Hypania invalida, Lumbricidae g.sp., Potamopyrgus antipodarum, Sphaerium rivicola and Pisidium spp. Thus the resulting saprobity in the Čunovo reservoir at Kalinkovo is worse beta-mezosaprobity. At Šamorín, two sites were also monitored. They were situated on a left-side and a right-side bay. Unlike Kalinkovo, the habitats were analogically represented by predominantly lenitic conditions with a larger depth (5-10 m) and muddy bottom. They were represented by a psammopelophilous zoocoenosis indicating already the alpha-mezosaprobity. The dominant species of this zoocoenosis indicating a worse degree of saprobity were Hypania invalida, Pisidium spp., Sphaerium rivicola, Chironomus spp. and Phytochironomus fodiens, as well as unidentified Lumbricidae g.sp.a Tubificidae g.sp. A survey of the recorded macrozoobenthos species and the degree of their abundance at individual profiles during the monitored period is presented in Tab. 1. (Tab 1 is situated as Fig. 6.) Conclusions Based on data obtained during monitoring of the qualitative and quantitative composition of macrozoobenthos and its relationship to saprobity in 2002-2005 at 8 profiles in the Danube between Bratislava and Medveďov, in the Čunovo reservoir and at beginning and end of the arm system, we can draw the following conclusions:
Proposals In regard to the fact that monitoring benthic evertebrates has become one of the important biological indices established by the new legislation for monitoring surface water in Slovakia and in the EU, I propose, in the interest of producing a complex evaluation of the Danube, expanding the network of profiles monitored by those situated downstream from Medveďov, downstream from the mouthing of the Váh river into the Danube at Komárno, in Radvaň nad Dunajom, downstream of Štúrovo, and possibly also at the mouthing of the Morava river. Furthermore, it is necessary in the interest of respecting new observations to revise the norm STN 83 0532 with comments, including a list of indication organisms and their modified indicative values. References
V.2.3. Long-term changes in the community of planktonic crustaceans, cladocerans and copepods in monitored localities in the Danube within-dike zoneMarta Illyová Bioindicative abilities of planktonik crustaceans Cladocerans and copepods represent a very important component of zooplankton of standing waters. They have optimal conditions in lenitic water bodies, in which the zooplankton is autochtonous, hence it forms directly in the given locality. On the contrary, flowing waters are unsuitable for development of zooplankton and relatively poor communities (dominated by rotatorians) form only in downstream stretches of lowland rivers. At higher water levels the planktonic crustaceans drift into the main stream from the side arms [10] Hence, they are allochtonous here and enrich potamoplankton with true planktonts. In the free water, medial zone, euplanktonic species of the genera Daphnia, Bosmina, Moina, Eudiaptomus, Thermocyclops etc. predominate. These so-called true planktonts are significant filtrators of phytoplankton in the water bodies. In the littoral zone, a community of littoral (phytophilous and benthic) cladocerans and copepods arises. Their species diversity increases especially in places with rich macrovegetation, where the community is enriched especially by phytophilous representatives of the Chydoridae family, as well as the genera Ceriodaphnia, Simocephalus, Scapholeberis, Eucyclops, Macrocyclops etc. If the macrovegetation is richly developed in a water habitat, the littoral species can also spread into the medial. In this case we characterise them as tychoplanktonic, i.e. false planktonts. In a balanced habitat with stable hydrological regime the euplanktonic species prevail over the tychoplanktonic species in the medial zone. Any shift in the ratio of these ecological groups indicates changes in the dynamics of hydrological regime in the given habitat. Communities of planktonic crustaceans react to changes in the hydrological regime both from a short-term and a long-term perspective and so they are a good indicator of environmental changes. Within the framework of the project “Biomonitoring of the impact of the Gabčíkovo hydraulic structures on the natural environment” we monitor species composition, dominance and species diversity of taxocoenoses of cladocerans (Crustacea, Cladocera) and copepods (Crustacea, Copepoda) of the medial and littoral plankton at defined localities on the Slovak-Hungarian stretch of the Danube during a 14-year period. Results of this monitoring have been published in several papers [for example: 2, 11, 12, 3]. Methods of sampling and interpretations of results Collecting and processing samples This contribution is based on our 14-year results (1991-2004) of monitoring cladocerans and is completed by my three-year monitoring of Copepods (2003-2005). Samples of cladocerans and copepods were taken from the medial and littoral three times a year, so that they represent the spring, summer and autumn aspect. The samples were taken using a planktonic net with mesh size of 60 – 70 µm and conserved by formaldehyde. Cladocerans and copepods were counted in complete or proportional samples under microscope. PCA analysis was carried out by means of the Canoco program [8] in order to determine the relation of cladocerans to certain habitat types. The primary data included 64 species from 18 relevées (six sampling sites x three sampling periods). Analysis was based on transformed values of the relative abundance of individual species. Copepods were omitted in this analysis. Monitored sites - The Danube main stream (since 1992 the old main stream) profile at Dunajské kriviny (rkm 1840,5) profile at Gabčíkovo (rkm 1819,5) - Parapotamal type arms (since the damming partly discharging at Čunovo) the Bodícke rameno arm (rkm 1830) the Istragovské rameno arm (rkm 1815,5) - Pleisopotamal type arms the arm in Kráľovská lúka at Trstená na Ostrove (rkm 1825) the arm in Sporná sihoť at Kľúčovec (rkm 1804) the arm at Čičov (rkm 1800) – monitored since 1999 Influence of the Gabčíkovo hydraulic structures on communities of planktonic crustaceans Eupotamal In the Dobrohošť–Sap stretch of the Danube, the proportion of euplanktonic crustaceans (e.g. Bosmina longirostris, Daphnia galeata, Daphnia cucullata, Eudiaptomus gracilis, Cyclops vicinus) decreased after the Danube damming. This decrease was caused by reduced development of the plankton in the arm systems and, as a consequence, by its limited drifting into the old main channel. The data available show that representation of the true planktonts decreased over the subsequent ten years, while the tychoplanktonic species attained an equal or even higher quantitative proportion. Among cladocerans the benthic species of the genus Alona and Macrothrix hirsuticornis and Chydorus sphaericus and among the copepods Nitocra hibernica and Eucyclops serrulatus occurred here. The mutual proportion of both ecological groups considerably fluctuated during the monitored period. However, in the last five years the true planktonts, especially species of the genus Daphnia and Bosmina longirostris, B. longispina and B. coregoni, re-appeared in the upstream stretch at Dunajské kriviny as well as at Gabčíkovo. However, these cladocerans were not drifted from the side arms as in the past. We explain the presence of these species by a considerably slowing down of the stream at some places in this part of the Danube, which creates more suitable conditions for the development of planktonic community. As this stretch is discharging, the cladocerans occur only sporadically and we did not observe any richer populations of planktonts. Based on the species composition of cladocerans, we can postulate that the character of the former main stream (eupotamal) has only partially approximated the parapotamal, because the so-called stagnant period, occurring in the past during periods of low water levels, is now absent. Abandoning the concept of the canalised old main stream Danube and restoring an anastomozing river pattern (stream, branching and rejoining irregularly to produce a net-like pattern) [6] would surely be better also for the planktonic crustaceans than the present straight-line like and continuously discharging Old Danube stream. The connection of the main arms of individual arm systems would ensure the preservation of different water bodies, making development of planktonic crustacean taxocoenoses possible. Parapotamal type arms The changes that have occurred in this type of water bodies can be demonstrated by the example of the Bodícke rameno arm, which is part of a permanently discharging system. After the Danube damming, the abundance and biomass of zooplankton considerably decreased here, euplanktonic cladocerans and copepods were eliminated and the dominance of tychoplanktonic species increased. Gradually, in the course of the last ten years, the true planktonts (Bosmina longirostris, Daphnia longispina and D. cucullata) completely disappeared from this arm as a consequence of the permanently discharging regime. A community of littoral species very poor in the number of individuals and consisting of the cladocerans Alona spp. and Chydorus sphaericus and the copepode Nitocra hibernica has occurred in the medial already since 1995. It is also a consequence of the fact that water flowing into the arm from the more upstream arms is poor in nutrients, as the sedimen in more upstream stretches. For that reason, under existing conditions we do not expect a change in the crustacean planktons of this arm and such a change is not even necessary because the development of zooplankton is the consequence of the stagnant water regime, which would not be suitable for the arm system under existing conditions. In the Istragovské rameno arm, which has a predominantly stagnant water regime, relatively rich communities of planktonic crustaceans remained. They are dominated by euplanktonic species, especially of copepods. In the course of ten years we could observe a dominance of euplanktonic cladocerans (Bosmina longirostris and Diaphanosoma orghidani) and copepods (Cyclops vicinus, Acanthocyclops or Thermocyclops oithonoides). The situation in the arm started to change in recent years, when the abundance and dominance of tychoplanktonic species increased in the medial. There we have witnessed a rise especially of the phytophilous species of the genera Simocephalus and Pleuroxus. Chydorus sphaericus also achieved greater dominance. This is a consequence of the considerable decline in the water level, the gradual shallowing and silting of the arm and an increase of littoral vegetation. These changes indicate isolation of the arm from the within-dike zone, which results in an increase of littoral cladocerans bound to macrophytes. A shift in the community of cladocerans is also illustrated in Fig. 1. The concept of a new eupotamal, i.e. the creation of a new stream by interconnecting the main arms of individual arm systems, would be optimal for both these arms. This would ensure the preservation of various water bodies in the original within-dike zone, as suggested for example by Lisický [5] and Šporka [7]. Pleisopotamal type arms In the last ten years, the dominance of euplanktonic species has gradually decreased and the predominance of tychoplanktonic cladocerans and copepods increased in all not discharging arms in Sporná sihoť, Horný les and Kráľovská lúka. This process was slowest in the arm in kráľovská lúka, where especially euplanktonic copepods (Eudiaptomus gracilis, Cyclops vicinus and Acanthocyclops robustus) were considerably represented still in the first years after the Danube damming. However, approximately since 1995, a tendency of a rising percentage of tychoplanktonic species, not only of cladocerans, but also of copepods, was observed here, similarly as in Sporná sihoť. Significant changes were represented especially by an increase in the number of species of cladocerans and copepods. The number of species recorded is high and corresponds to species diversity of identical habitats of large European rivers [9, 4]. We have also recorded rare and sporadically occurring species of cladocerans like Latonura rectirostris and Bunops serricautata. Based on the classification of habitat naturalness (originality) [1] according to species number, these arms belong to the category of original habitats deserving protection. The increase in species diversity is also a consequence of the natural process of overgrowing, ageing and terrestrialisation of these water bodies, which are accelerated by simulated floods that have a lower frequency and intensity than natural floods did in the past. This process, however, leads to the inevitable extinction of these habitats and of the rich fauna of planktonic crustaceans in them. For this reason it is necessary to ensure regular communication of these water bodies with other water bodies in the within-dike zone, at least during the flood water levels, in order to enrich them by nutrients, to wash out sediments and to slow down process of overgrowing, shallowing and terrestrialisation. Changes in species composition of planktonic crustaceans During the course of monitoring we recorded 68 species of cladocerans and 48 species of copepods. We consider the recorded number of taxa to be high. At present we expect about 100 species of cladocerans to occur in Slovakia, among which as much as 68% occur in the seven sites monitored. During the last 14 years the species number [3] has increased especially for the following reasons: a) penetration of invasive species into the Danube floodplain (e.g. Pleuroxus denticulatus, Eurytemora velox, Ectinosoma (Helectinosoma) abrau); the species Disparalona hamata and Ilyocryptus cuneatus were found in Slovakia for the first time, b) long term monitoring also recorded sporadically occurring species, c) the development of littoral vegetation made an increase in the number of phytophilous species bound to macrovegetation possible. The largest increase in species number was recorded in the pleisopotamal type arms, which (according to Ward [13]) documents a low degree of communication of habitats in the floodplain. Evaluation of results of monitoring and its significance for the future Changes in hydrological regime of the territory affected by damming of the Danube main stream have significantly influenced the structure of taxocoenoses of cladocerans and copepods. Based on these changes we support the proposed measures for optimising the hydrological regime in this area. They should lead to the establishment of conditions that better approximate natural ones. It is necessary to continue monitoring planktonic crustacean communities in order to be able to evaluate whether the measures carried out will lead to the set goals. References
V.2.4. Benthic fauna in monitoring of biota in the area of the Gabčíkovo hydraulic structuresFerdinand Šporka In 1990-1996, the Polychaeta, Oligochaeta, Amphipoda, Isopoda, Mysidacea, Chironomids and Simuliidae were included in the monitored groups of aquatic animals. As this part of the monitoring was excluded from monitoring due to a lack of data from the Hungarian side, we evaluate only the initial stage of changes caused by the Gabčíkovo structures according to changes caused in selected groups of permanent fauna. We exclude, in this case, chironomid and simuliid larvae. All these groups reacted very sensitively both to the change in hydrological regime and the changes in the character of habitats in water bodies in the former main stream of the Danube (Dunajské kriviny, Istragov), in the discharging arms of the parapotamal type (Bodícke rameno, Istragovské rameno) as well as in temporarily discharging arms of the pleisopotamal type (Dunajské Kriviny, Kráľovská lúka). These changes were evaluated in detail in several papers [4, 2, 7, 3, 5, 6, 8], which produced data on the pre-dam composition of aquatic biota and changes in its composition after, two, six and ten years of monitoring. They state that stream velocity in the former main Danube stream decreased and that the gravel sediments stabilised as a consequence of reduced discharge in the upstream parts (intake place Dunajské kriviny). Under conditions of considerably shallower and more transparent water (sedimentation of fine particles in the Čunovo reservoir), an increase in algae occurred there. The algae increased the amount of food available for scrapers and shreders, especially for Amphipods and Isopods, and for species inhabiting and eating the algal growths (family Naididae, Oligochatea). Due to the increased food offer, the abundance and biomass of these groups increased several fold. Species atypical of such a habitat type appeared, such as species of the genus Limnodrilus (Tubificidae, Oligochaeta), which previously occurred only in the arms. Backwater appeared upstream of the mouthing of the tailrace canal into the Old Danube and, in the locality Istragov, caused a slowing down of stream velocity, and also evoked sedimentation of fine silt-like particles. Such sedimentation was observed especially in periods of increased outflow of water from the tailrace canal into the Danube. The original gravely bottom was covered by mud (layer of silty-sandy sediment), which caused a change in the structure of the original benthic community. In the profile of Istragov, the Amphipods that dominated there in the pre-dam period disappeared, while new representatives of Oligochaeta appeared, which had either not occurred there before or only occurred there to a limited extent. They were represented by species of the genus Limnodrilus bound to the muddy substrates. Their abundance gradually increased from 350 ind.m-2 up to 40 000 ind.m-2. The documented changes in the former arms of the parapotamal type, which became discharging similarly to in the occasionally discharging of the pleisopotamal type, were published by Šporka & Nagy [8, 4]. The essential change in the discharging arm was their approximation to the benthic fauna of the Danube mainstream, hence colonization by the rheophilous species. The former occasionally discharging arms of the plesiopotamal type (Kráľovská lúka) are not washed by water to a degree sufficient to inhibit overgrowing by macrovegetation. In pre-dam conditions the water velocity in the arm approximated, under flood states, the water flow velocity in the main stream and effectively inhibited the development of water macrovegetation. What’s more, their supplying by surface water made possible the intake of biogenous elements (N, P), which are bound, in the case of an insufficient supply by surface water, in aquatic macrophytes (own unpublished data). That is why strong overgrowing and accelerated terrestrialisation has started in the type of arms represented by Kráľovská lúka, with species bound trophicaly to macrophytes predominating there. The largest change was observed in the former arm at Dunajské kriviny. This arm is situated upstream of the intake structure canal and is not supplied with water. The arm was without water almost all year over the period 1993-1997. Water appeared in it only in the spring and at the deepest places. On the contrary, improved conditions for benthic fauna appeared in the Žofín arm, where modifications of the hydraulic regime made the permanent supplying with water from surrounding water bodies possible. Proposals for improving common monitoring Proposals of changes to and supplementing of monitoring, monitored area, localities and monitoring methods The existing mode of sampling, processing and evaluating the monitoring of aquatic biota, especially benthic fauna, no longer satisfies the requirements made of modern monitoring. The Water Framework Directive Directive (WFD) 2000/60/EC requires the complex monitoring of the entire benthic fauna – aquatic invertebrates. Upon assessment of the ecological state of water bodies, autecological characteristics of macroinvertebrates are used, based on which indices – metrics – are acquired, which make their classification possible. Until 1997 the macroinvertebrates were monitored in complexity, then the sampling and collecting of all groups was desisted, with only some groups being monitored at present. From this perspective the current range of groups of monitored hydrobionts is insufficient. The WFD for assessment of the ecological condition of water bodies is based on four main groups of water biota (phytobenthos – phytoplankton, macrophytes, macroinvertebrates and fish). Zooplankton is not included in the directive, but it’s monitoring just in the arm systems must be continued in any case. Accounting for the WFD, it is also necessary to modify the mode of sampling and the processing of samples so that sampling and sample processing are executed in uniform manner, with their identification being carried out by individual specialists only subsequently. In this way we achieve a situation where macroinvertebrates from one sampling site are evaluated, which was not provided to date. Individual specialists take samples from different habitats, which make the comparison of changes impossible. For example, samples of zooplankton and fish are taken from the remnant of the arm at Sporná sihoť, while other monitored groups of aquatic insects are taken from the Opátske rameno arm (Bulánková – Krno in this issue), which represent a different type of aquatic habitat. Proposal of new method of sampling macroinvertebrates. As mentioned above, the monitoring of macroinvertebrates should be based on the taking of one common sample, whereby after sorting individual taxonomic groups are to be given to respective specialists for identification. Sampling should be modified, on both the Slovak and Hungarian sides, in accordance with methods required by WFD. As sampling in the medial zone of the former mainstream and arm requires expensive sampling devices, I suggest taking samples for the purposes of monitoring only in the ripal or littoral zone according to methods proposed by the consortium AQEM and STAR. In this mode a hand net (kick net) is used and a total of 20 samples are taken, each when calculated comprises a surface area of 1.25 m2 from different sediments. The estimation of sediments is carried out on a stretch of 100 m so that a representative sample is obtained. This mode of sampling has been published in the methodical directive proposed by the AQEM Consortium [1]. The same mode of sampling has already been used in Slovakia in sampling from reference and monitoring sites for the purposes of the WFD. Proposal of common monitoring of natural environment in the stretch between Sap and Štúrovo The network of existing sampling sites and any proposals for changes and the supplementing by new sites for monitoring the stretch Sap – Ipeľ river mouthing – Budapest, should be subjected to deeper analysis in a joint working group of specialists. References
V.2.5. Use of terrestrial molluscs for bioindication of the impact of the Gabčíkovo hydraulic structuresTomáš ČejkaBioindication abilities of terrestrial molluscs The advantages of molluscs as a model group can be characterized as follows:
Calibration of indication properties It is known that animals, hence also molluscs, can often have relatively different ecological requirements in different zoogeographical units. Thus, knowledge of habitat requirements of the iuxta-Danubian (close, at, beside the Danube) molluscs cannot be automatically applied for example to conditions of the Borská nížina lowland and not at all to more remote parts of Europe under the influence, for example, of the Atlantic climate. For this reason, results of monitoring from geographically and climatically distant areas are not fully comparable. Prior to a monitoring investigation, a calibrating indication investigation must be carried out in each area where the local ecological requirements of individual species are not known in detail in order to determine (calibrate) them by means of the methods given below. A brief description of methods and interpretation of results Field and laboratory work When evaluating changes in the natural environment by means of mollusc communities, the quantitative and semi-quantitative methods are used. The most applied method is the volume method, with which a standardised volume (mostly 5 litres) of litter and soil upper layers are taken from several places in a homogenous plot, approximately up to a depth of 3-5 cm. After washing, drying and subsequent sieving, the mollusc conches are sorted from individual fractions, identified and counted. The proportion of individual species in a sample of the community determines its character (mutual proportion of hygrophilous, shadow-preferring, hygrotolerant or euryecious species). Another, less frequently used method is the square method, which differs just by collecting litter and soil substrate over a specific surface area (e.g. 50x50 cm). In this case the number of molluscs is related to the area unit. Analysis and synthesis of data Counting individuals in samples gives us absolute data, which should firstly be transformed (converted to a scale determined by experts, transformed logarithmically, etc.), to be prepared for statistical analysis. In addition to usual descriptive quantitative characteristics like population density, relative abundance (dominance) and structural characteristics like diversity and equitability, data can be subjected also to different methods of multivariate analysis (cluster analysis, ordination), which helps us visualise the data structure and makes data interpretation easier. Results obtained in individual years can be continuously compared and the trends estimated. Principles for selection of investigation plots and problems of data interpretation Within the scope of the monitoring plots set by the monitoring coordinator, the structurally most homogeneous parts are to be selected, in which specific investigation plots are then established. Investigation plots of 50 x 50 m are suitable. The investigation plots should be situated as far from the stand margin as possible to eliminate the edge effects. It is important to ensure that the monitoring plots remain unaffected by silvicultural interventions (e. g. thinning or even timber exploitation) during the whole period of monitoring. Otherwise, we would be monitor the influence of silvicultural, but not the influence of changes to hydrological conditions. Problems with sampling arise on those plots that are flooded by surface water several times a year. After a flood most molluscs are washed away from the plot or the local patchy structure of communities is mixed by water turbulence. For this reason the samples obtained 5-8 weeks after the flood are hardly interpretable from the viewpoint of moisture changes. Another problem of data interpretation arises in the habitats situated at a considerable distance from the flood zone and which remain for a long time out of the influence of floods, especially in habitats situated on elevated places (terrace platforms, fluvial levees), where the ground water table is deep and does not influence the hydropedological regime in the soil upper layers. In such a habitat it is mostly problematic to interpret changes in mollusc communities as a direct consequence of water regime changes. Influence of changes to surface and ground water dynamics on communities of terrestrial molluscsMolluscs, and their communities, reliably react to changes in soil moisture and the absence or presence of floods. (a) Habitats frequently flooded by more or less destructive (streaming) surface floods are characterised by the exclusive presence of polyhygrophilous species and by a naturally low alpha-diversity of communities (in average 12 species). The poorest in the number of species are mollusc communities in the initial stages of floodplain forest (in average 6 species) [3, 4]. Only those species that are adapted to flood conditions survive there. They are represented especially by larger species that are able to escape from the rising water level by climbing onto trees (Arianta arbustorum, Cepaea hortensis, Succinea putris, Oxyloma elegans, Trichia hispida, Trichia striolata danubialis) or that are able to survive for a time directly in the water or in different water-bearing substrates, especially in deposits of plant material (Zonitoides nitidus, Pseudotrichia rubiginosa, Carychium minimum, and partly also probably Trichia striolata danubialis and Trichia hispida). In recently flooded habitats, which are covered after an intensive flood by a silt crust, the polyhygrophilous species Oxyloma elegans, Succinea putris, Zonitoides nitidus and Pseudotrichia rubiginosa appear as pioneer species. Other strongly hygrophilous species, Carychium minimum, appears later, because it is a small species having a small dispersal power, and the oligotrophic type of substrate does not satisfy its trophic requirements. (b) In sporadically flooded parts of the floodplain, which are influenced by surface waters only at high or very high water levels in the Danube, but which have a high level of ground water and a moist to waterlogged substrate, the dominant species are Carychium minimum and Zonitoides nitidus, while Succinea putris, Arianta arbustorum, Trichia striolata danubialis, Trichia hispida, Deroceras laeve or also Euconulus praticola occur as subdominant species. (c) Habitats with the natural or semi-natural hydrological regime mentioned above, which see a sudden drop in the ground water level or the halting of the influence of floods, are at first penetrated by moderately hygrophilous species (Semilimax semilimax, Cochlicopa lubrica, Succinella oblonga, Vitrea crystallina), while the polyhygrophilous species start to decline and free up niches for mesohygrophilous or eurytopic forest species (Alinda biplicata, Monachoides incarnatus, Fruticicola fruticum, Aegopinella nitens) or even for euryecious or anthropotolerant species (Punctum pygmaeum, Limax maximus, Helix pomatia). According to the degree of moisture the new community gradually obtains a structure corresponding to a specifically drier habitat type, mostly characteristic of a degraded community with a certain portion of xenocenous and anthropotolerant species (Limax maximus, Arion fasciatus, Arion lusitanicus). Influence of silviculture on molluscs From the 48 species of terrestrial molluscs regularly occurring in the iuxta-Danubian floodplain forests, as many as 32 species also occur in older poplar lignicultures [1, 3]. According to existing data, the most constant species inhabiting the stands of poplar cultivars are Monachoides incarnatus, an eurytopic forest species, which reaches a dominant to subdominant position here, and Fruticicola fruticum having similar ecological requirements as Monachoides incarnatus, but only managing a subdominant to receding position here. Among the constant species, the forest mesohygrophilous Aegopinella nitens, forest hygrophilous Arianta arbustorum and forest eurytopic Alinda biplicata also should be named. Cochlicopa lubrica, Cepaea hortensis and Trichia striolata, having optimum conditions in moister sites in willow-poplar forests, and forest hygrophilous species Vitrea crystallina, Urticicola umbrosus and Clausilia pumila, as well as forest mesohygrophilous Cochlodina laminata occur here mostly as subdominant species [3]. Richterová [11] found 28 species in poplar ligniculture of the within-dike zone. The most frequent were forest hygrophilous species Arianta arbustorum, Trichia striolata danubialis, Vitrea crystallina, Cepaea hortensis and Fruticicola fruticum, mesohygrophhilous forest species (Aegopinella nitens, Monachoides incarnatus) and polyhygrophilous Carychium minimum. As dominate species she gives Aegopinella nitens, Punctum pygameum and Carychium minimum, but the largest portion of the community consisted of euryecious species (22%) and eurytopic forests species (17%). The above enumeration shows that the forest species of the Illyrian regiotype (sensu Lisický [6]) were able to successfully adapt to conditions of economic forest stands and to cyclical anthropogenous disturbances (clear-cuttings, overall soil preparation etc.). Even species of the regiotype of forest permanent dwellers do not represent an exception. The poplar cultivars have their moisture optimum in dry types of softwood floodplain forests (especially at sites of the Salici-Populetum typicum variety with Swida sanguinea), in transitional floodplain forests (Fraxino-Populetum) and in the surroundings of Bratislava also in sites of moister hardwood floodplain forests (Ulmo-Fraxineta). Hence, individual habitat types have a corresponding structure of mollusc communities. We do not exclude the potential occurrence of Cochlicopa nitens, Succinella oblonga and Euconulus praticola in moist habitats. They are present in this territory, but occur rarely. As to Columella edentula, to date it seems that it escapes medium-moist habitats (transitional floodplain forests and drier varieties of willow-poplar floodplain forests) and very moist habitats (it probably cannot tolerate floods) [3]. In young stands of poplar cultivars, in dependence on the habitat, usually Trichia hispida (in moister habitats) Aegopinella nitens, Monachoides incarnatus and Urticicola umbrosus (in drier habitats) predominate. Remarkable is also the occurrence of other forest species, for example Cochlodina laminata and Trichia striolata, whose occurrence in this type of stands was not originally expected. Insufficient shadowing by the tree canopy in the case of young stands is substituted by high and dense ruderal vegetation and, if stumps remained at the place of clear-cut plots, also “sprout enclaves”, in which even some forest hygrophilous species are able to survive due to a favourable microclimate. Long-term changes in the structure of iuxta-Danubian fauna Structure of iuxta-Danubian mollusc fauna in the past (evaluated based on thanatocoenoses) A knowledge of the history of development of hydrological and habitat conditions in individual plots is necessary foremost in order to produce an objective evaluation of the so-called zero state of the natural environment, but also for evaluation of the subsequent sequence of changes caused by the changed hydrological regime. In spite of existing local investigations, the general question still remains as to when and how the present iuxta-Danubian geobiocoenoses originated? There is a relatively clear answer in the case of floodplain forests, especially for the strip of floodplain forests in the within-dike zone. The floodplain forests doubtlessly existed there already in the prehistoric period (younger Holocene) [5]. They were, however, probably much more extensive, and so the present strip in the within-dike zone is just a reduced relict of them. In the Danube calciferous sediments, mollusc conches preserve well and often form complete thanatocoenoses (death assemblages). According to thanatocoenoses, which can be dated, in addition to radiocarbon methods, also on the basis of old maps, the habitat conditions of specific localities can be relatively accurately reconstructed. Mostly a complete successive series of the habitat can be reconstructed. According to existing findings of Holocene’s mollusc fauna, it is probable that a border separating the wide zone of flooded floodplain forests from the elevated, drier and partly steppe-like interior of the Žitný Ostrov Island run approximately along the area of the present village of Vrakúň. Hence in the past (in younger Holocene), the floodplain on the present Slovak territory had a width of about 10 km (today in the widest part at the village of Baka it is only 3 km) [8]. The question of the steppe-like enclaves within the floodplain is dealt with in [8, 9]. As yet the dating of individual processes is missing, together with a knowledge of the topographical distribution of individual geobiocoenoses, which can be made more accurate only by further investigation of recent and fossil mollusc fauna, prehistoric settlements and by searching for data in archive documents from this area. From the perspective of the changes caused by the Gabčíkovo hydraulic structures, the period before the so-called Lanfranconi regulation of the Danube for the medium water (1886-1896) is of particular interest. The mollusc thanatocoenoses in the iuxta-Danubian area were studied by Pišút & Čejka [10]. According to historic maps, they dated the absolute age of sediments and by means of mollusc thanatocoenoses reconstructed the complete succession series of the floodplain forest from its initial stage to the present transitional floodplain forest (period of 1791–1999, i.e. 208 years). They confirmed the occurrence of similar types of floodplain forests as those occurring in the iuxta-Danubian area at present and the occurrence of thanatocoenoses, whose structure approximately corresponds to the structure of the most preserved forest mollusc communities occurring at present in the area between the villages Dobrohošť and Sap. The total area of the moistest types of softwood floodplain forests with corresponding malacocoenoses was, of course, much larger in the past than at present and showed a more natural structure of stands. This is obvious even upon comparison of phytocoenological relevés from the 1940-1950-s with the present ones [12, 13]. Unfortunately, there are no quantitative or semi-quantitative data available on mollusc communities in the iuxta-Danubian area from the first half of the 20th century. Therefore the only possibility of comparison is an extensive paleoecological investigation of recent Danube sediments. Mollusc fauna structure before the Danube damming and ten years on. In the beginning it must be noted that before the Danube damming there were two principal types of mollusc communities: (1) mollusc communities of the softwood, willow poplar floodplain forests and (2) those of hardwood floodplain forests. The iuxta-Danubian area can be divided from the viewpoint of mollusc communities into the following parts: (1) the surroundings of the Čunovo reservoir on the stretch Bratislava Dobrohošť, (2) the by-passed area of the Gabčíkovo hydraulic structures on the stretch Dobrohošť – Sap, (3) the stretch downstream from the mouthing of the tail-race canal (Sap – Klúčovec), (4) the area of Čičov village out of the direct influence of the Gabčíkovo hydraulic structures. In the first stretch, the level of ground water increased after the Čunovo reservoir was filled and the hydropedological regime became renaturalized. The habitats in this area were, however, void of any larger influence of the groundwater level and relied exclusively on rainwater. The original hygrophilous communities disappeared and the habitats in the course of revitalization are lacking in immigration sources of autochtonous mollusc populations due to the absence of the possibility of passive (drifting by floods) or active spreading [2, 3]. In the second stretch, after the damming, the groundwater level declined in the zone running along the Danube old main channel. The changes appeared first of all in the representation of individual subtypes of softwood floodplain forests with characteristic structure of mollusc communities. In the by-passed area molluscan communities of the moistest types of floodplain forest are disappearing, especially those bound to the subassociations Salici-Populetum phragmito-caricetosum and Salici-Populetum myosotidetosum. After the damming, most of these types of mollusc communities started to restructure on “drier” types bound to various degraded types of willow-poplar floodplain forests (Salici-Populetum typicum). In the third stretch, the Danube riverbed bottom deepened, but the water levels were not significantly changed. For the moment this has not reflected on the communities of terrestrial molluscs, but in future we cannot exclude adaptive changes caused secondarily by vegetation changes resulting from the eventual changing of the ground water level. The fourth stretch at the village of Čičov is out of the direct influence of the Gabčíkovo hydraulic structures. Expected development of the mollusc communities structure while saving the present hydrological regime. The existing hydrological regime is relatively satisfying only from the viewpoint of revitalization of ground water levels at the Čunovo reservoir (the floodplain stretch downstream from Bratislava). However, with some exceptions, this area is not regularly flooded. Upon high discharges in the Danube, some parts of the area in the stretch between the Prístavný Most Bridge and Čunovo are flooded (for example at the Rusovské Ostrovy Islands). Areas situated behind the flood protective dikes are not flooded. Accounting for the relatively high level of ground water, the conditions for terrestrial mollusc are potentially satisfying. However, the polyhygrophilous species have almost disappeared due to insufficient moisture and the absence of floods. These species are already unable to colonize these habitats either passively or actively. Some chance for the revitalisation of mollusc communities might be the relocation of polyhygrophilous species from areas situated more downstream (Kráľovská lúka, Erčéd). A worse situation exists on the stretch Dobrohošť – Sap, where the absence of natural floods and results in negative structural changes in the mollusc communities. The floods can be partly simulated by leading water into the arm system from the intake structure at Dobrohošť, but there is not a sufficient will to carry out such flooding because of conflicts of interests. The disadvantages of simulated floods are dealt with in the monograph [7], which presents ideas about the revitalisation of the area. The present floodless situation is bad and the succession of most of the originally strongly moist habitats will stop in the stage of dry types of willows poplar floodplain forests, or even drier succession stages, which will concurrently influence the succession of respective mollusc communities. References
|
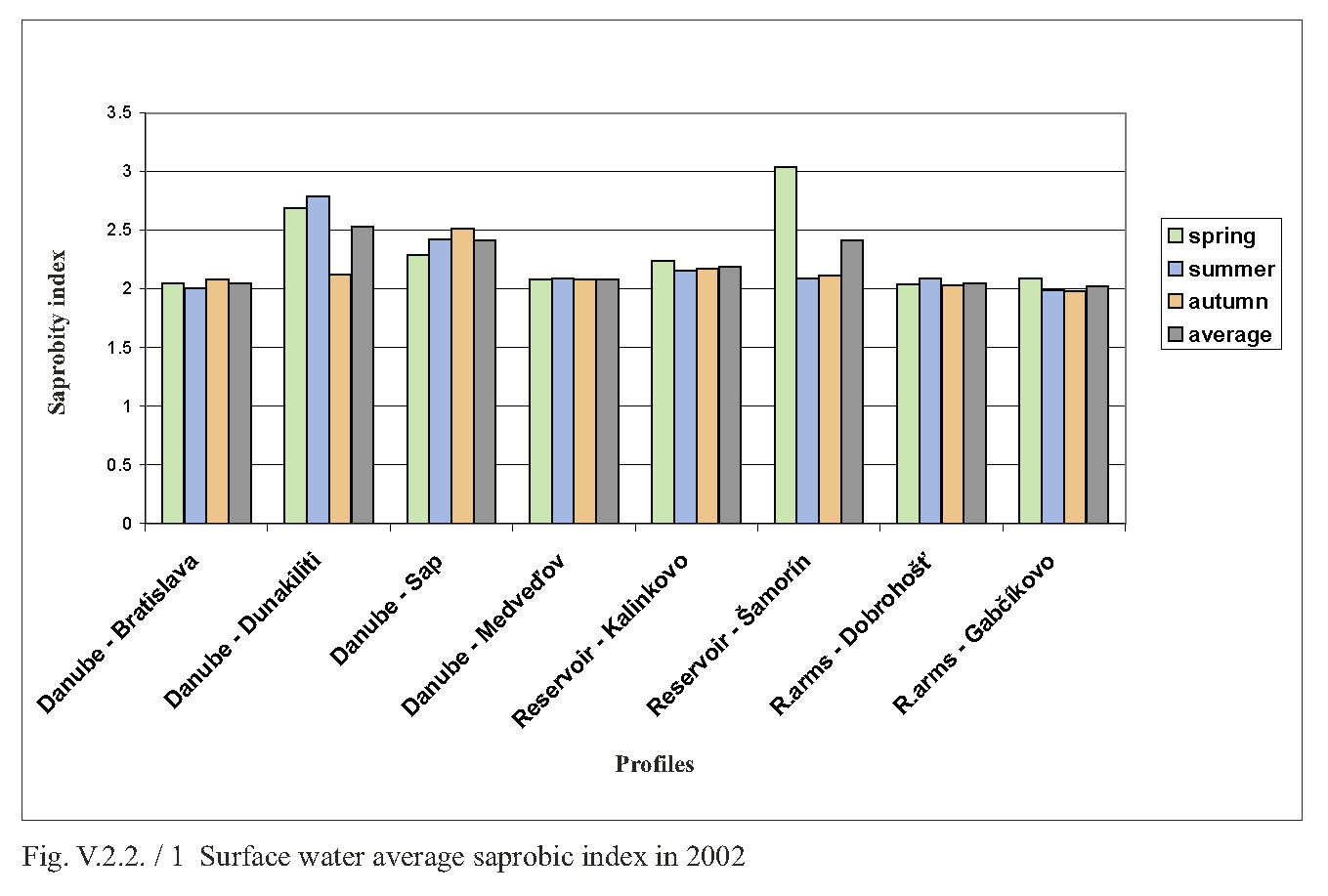{kind=link}
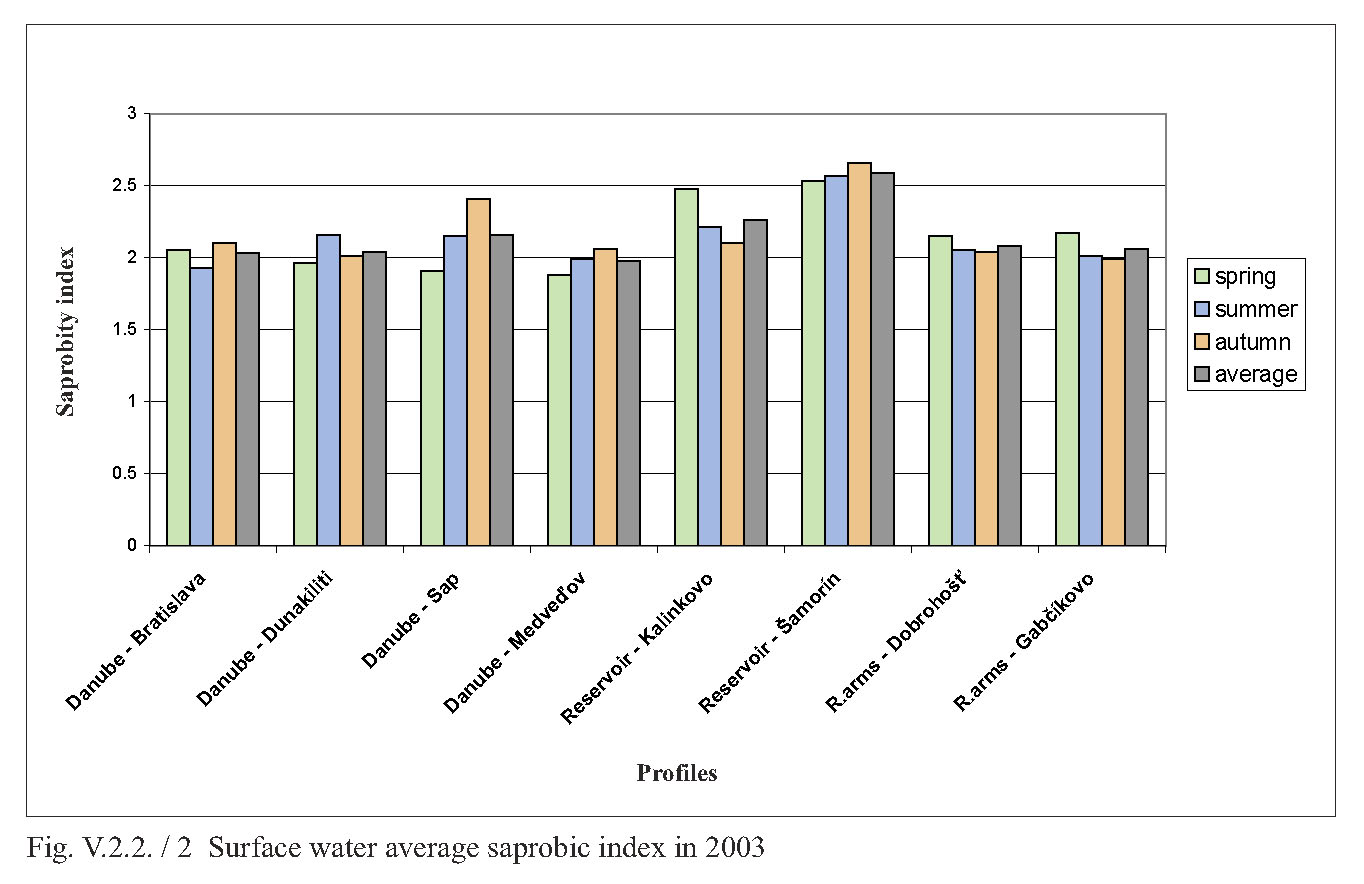{kind=link}
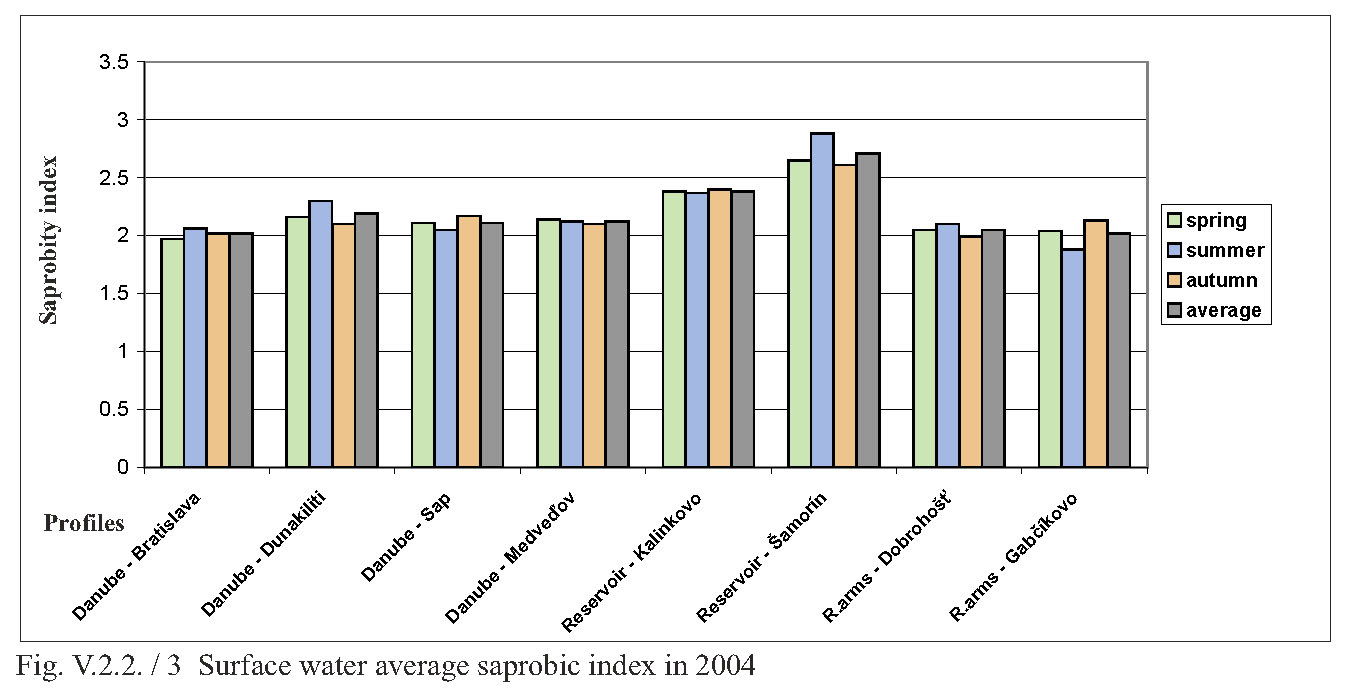{kind=link}
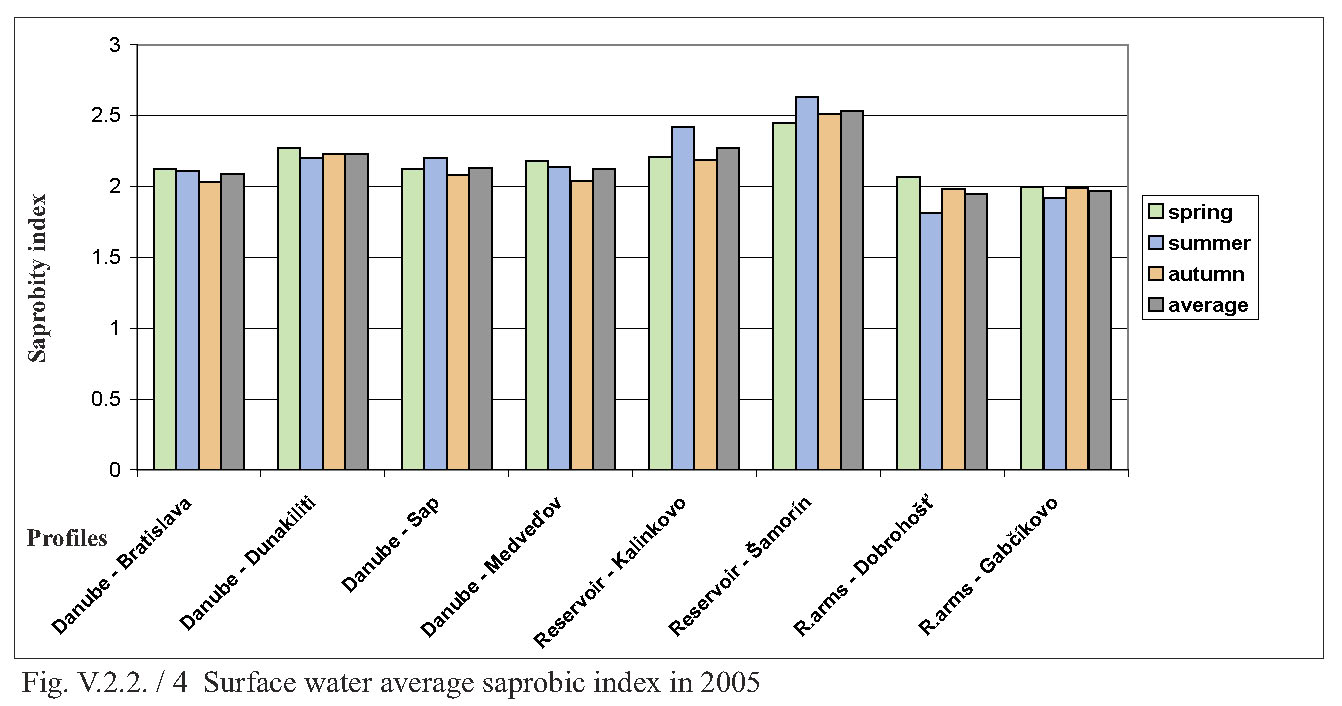{kind=link}
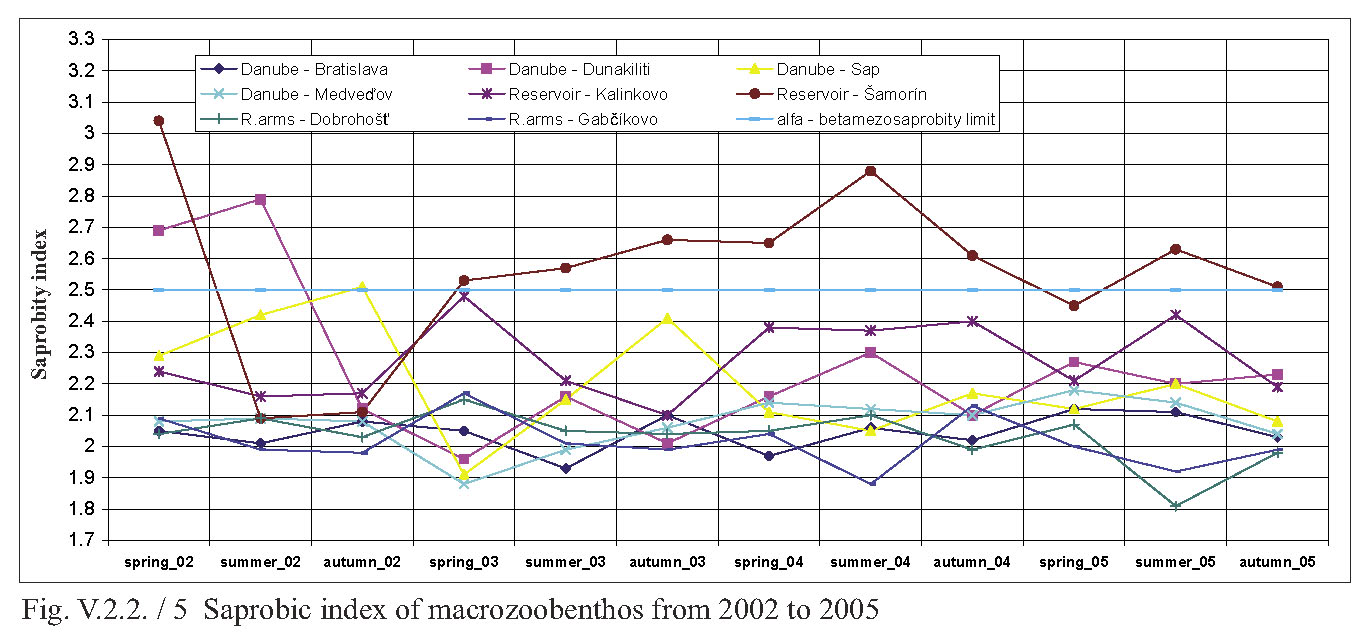{kind=link}
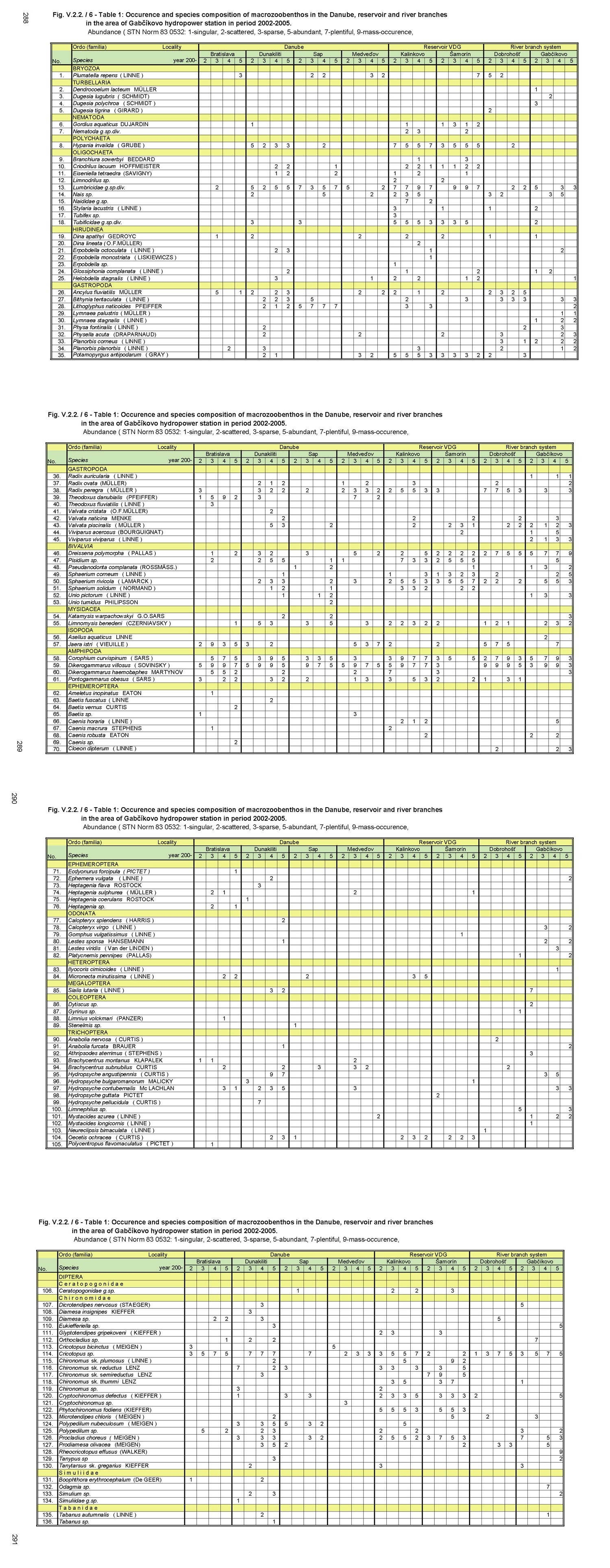{kind=link}
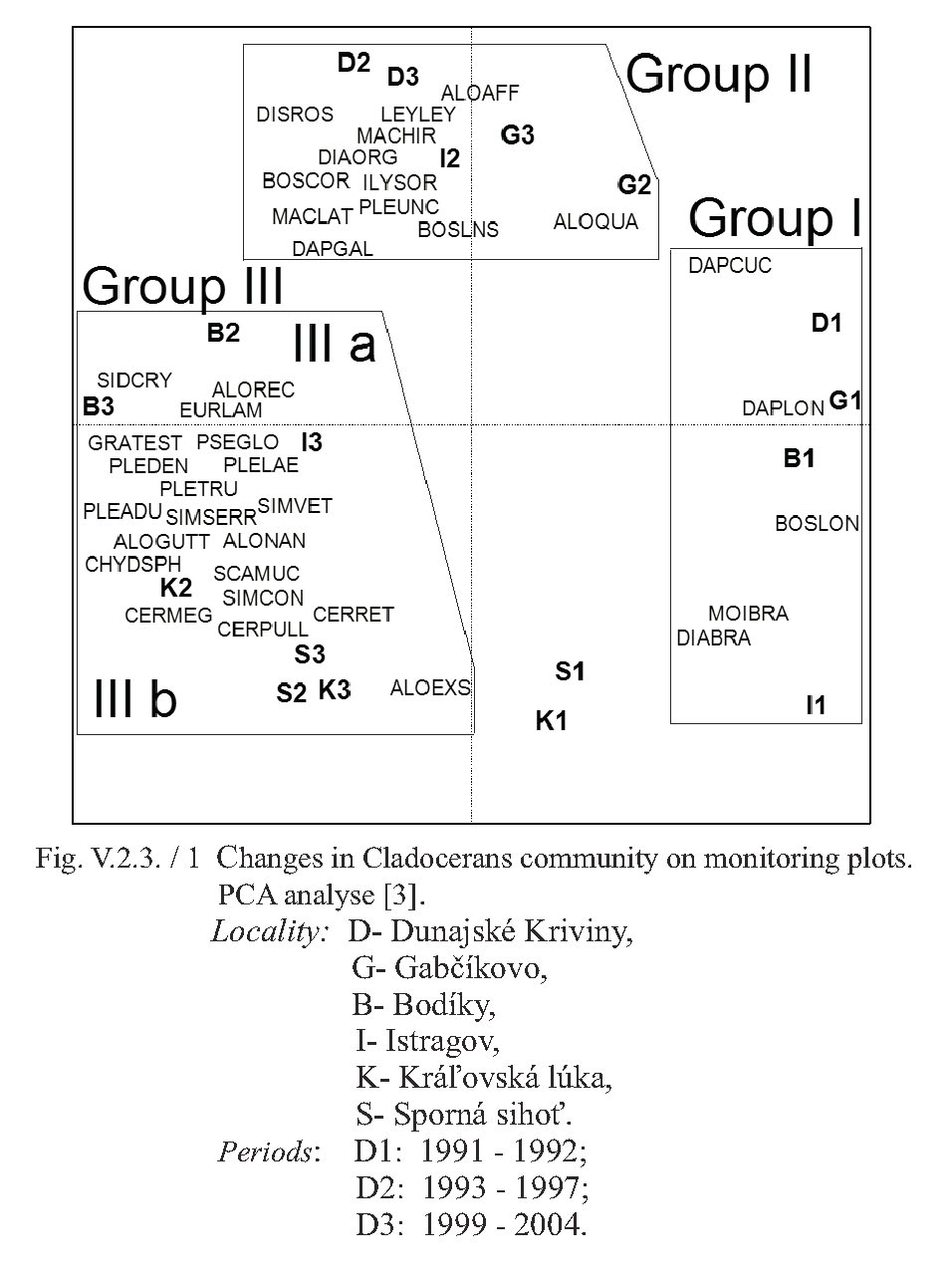{kind=link}