CONCLUSIONS Changes in weevil taxocoenoses are dependent upon a set of factors of exogenous as well as endogenous importance. For a relevant evaluation of the impact of the Hydroelectric power structure Gabcikovo on a part of the edaphon, a complex study using precise quantitative method is necessary. Up until the present results of study of weevil taxocoenoses and their changes cannot be evaluated as an impact of the Hydroelectric power structure Gabcikovo. Only further studies will show more precisely the interpretation of the relation between the taxocoenose, changes and the impact.
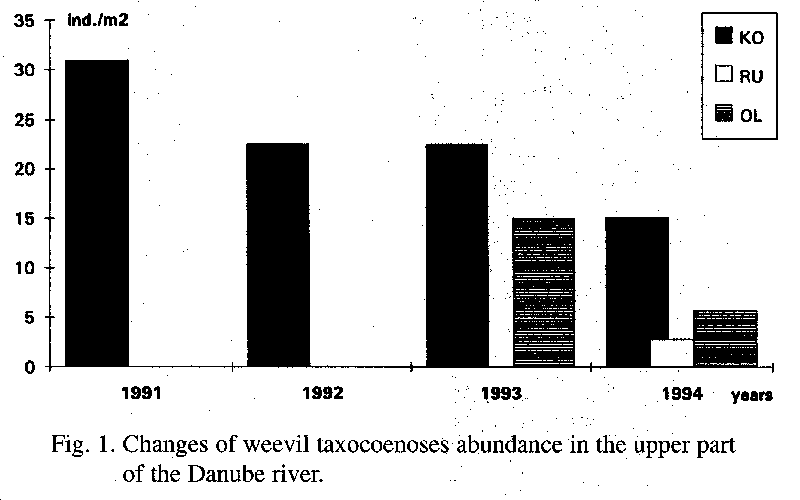
It is important to record the influence of exogenous environmental factors for monitoring. The following factors were observed: pH value, contents of CaCO3, river floods and, extreme summer droughts. Forest stand structure has an influence only in the composition of the following weevil taxocoenoses (Lignyodes sp., Stereonychus sp., Byctisus sp., Tanymecus sp., Curculio sp.). The Danubian floods are a decisive phenomenon. The flood in the 1991 manifested itself most decisively. The abundance of weevils after the flood was low in each area (Fig. 1, 2, 3), because the soil humus layer was affected during the flood.
Weevils are phytophagous beetles settled in various biotopes. In the soil habitat they occur either accidentally (geoxenes), or occasionally (geophiles), or their whole development takes place in this habitat (geobionts).
Data on the changes of weevil taxocoenoses have been published in papers [1-7].
Based on the data on the abundance, dominance and bionomy I also described the types of socions for some areas. The type of socion characterizes the dominant (geobiont or geophile) and the variant (species with maximal frequency of occurrence in the observed territory). For the areas KO, OL it is the Barypeithecium chevrolati variant with Acalles echinatus. For area DO it is the Barypeithecium chevrolati variant with Dorytomus puberulus. For area BO it is the Sciaphilicium asperati variant with Dorytomus nebulosus. For areas KU and CI it is the Dorytomicium puberuliti variant with Acalles echinatus. It was not possible to list the socions for other areas, until now.
Only two species, Dorytomus puberulus and Ceutorhynchus pallidactylus had a permanent distribution in the observed areas, during the four years. I also observed some rare species in the socions of weevils: Gronops inaequalis, Otiorhynchus velutinus, Trachyphloeus parallelus, Trachyphloeus asperatus, Bagous limosus, Grypus brunneirostris, Bradybatus creutzeri and Rutidosoma globulus.
The highest average abundance value of 32 ind.m-2 was recorded in the area KO in 1991. In 1994 the abundance dropped to half of the value in the first research year. Also at the same time the number of weevil species decreased from 28 to 12. In years 1992 and 1993 the abundance was 22.5 ind.m-2 and the number of species 32 and 31, respectively.
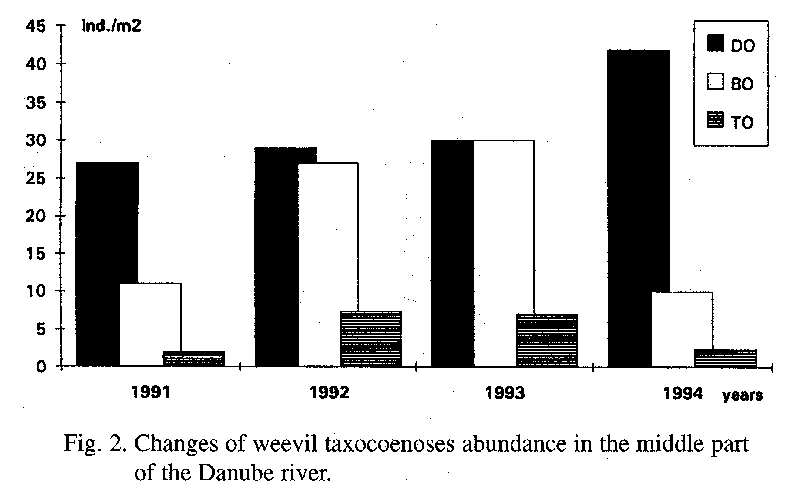
Since 1991 in the area DO the abundance had an increasing character, from 27 to 42 ind.m-2. The high abundance in the last year of research was caused by the species Barypeithes chevrolati. The maximal abundance of this species was 165 ind.m-2 in May 1994. The number of species in this area varied from 14 to 30.
In 1992 the maximal abundance in the area BO was 27 ind.m-2, and in 1993 30 ind.m-2, with the number of species being 25 and 24, respectively. The abundance was approximately the same in the other years with 11 and 10.6 ind.m-2, with the numbers of species being 29 and 22.
With a 2 to 7.4 ind.m-2 the abundance is small in the area of TO. In the first and the last year of the research the abundance of weevils was with a similar 2 and 2.4 ind.m-2 and the number of species was 12 and 8, respectively.
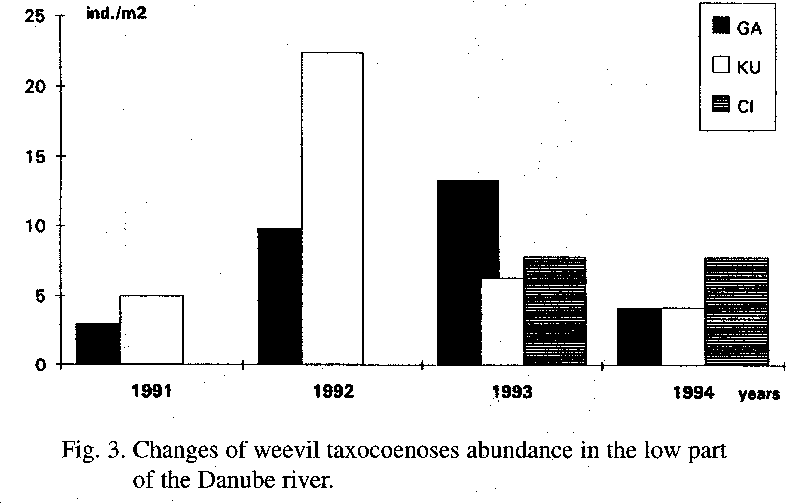
A similar course of abundance as in the areas BO and TO was observed in the area GA. In the first year 3 ind.m-2, and in the last year 4.2 ind.m-2, with the number of species being 12 and 14, were found. I observed maximal values in 1992 and 1993 (9.8 and 13.3 ind.m-2), with a corresponding number of species of 19 and 19.
The first and the last year is again similar in the abundance value in the area KU (1 and 4.2 ind.m-2, with the number of species being 14 and 14). The highest values were observed in 1992, with 22.4 ind.m-2, with the number of species being 30.
The areas OL and CI were observed for two years. In 1993 the abundance in the area OL was 15 ind.m-2, with the number of species being 26, and in 1994 5.7 ind.m-2 (15 species) were found. The abundance in the area CI was with 7.8 in.m-2 in 1994 the same but with a different number of species; 16 in 1993, and 8.
The area RU was observed only in 1994. The average abundance was 2.8 ind.m-2, with the number of species being 9.
In evaluating of the changes of the abundance dynamics of weevils the whole observed stretch can be divided into three parts. The upper part, Bratislava-Cunovo, lying at the reservoir including the localities of KO, RU, OL, the middle part with the localities of DO, BO, and TO which is situated between the by(pass canal and the old Danube river bed, and the lower part, downstream from the hydroelectric power plant including the localities of GA, KU, CI.
The upper part is not influenced by floods. In the area of KO the tendency of the lowering of the abundance dynamics of weevils can be observed. This tendency is caused mainly by the geophilous species of the genus Dorytomus. Their diminution may be dependent on the desiccation of the tree species Populus sp. An increase of the water level and the prepared water supply in the Biskupicke rameno river arm, will cause a certain "revitalisation" of the floodplain of Ostrov Kopac.
In the middle part the course of weevils abundance is similar in all areas (Fig. 2). The abundance values are different only in different years of the research. In 1994 the abundance increased in the area DO. This is caused only by the species Barypeithes chevrolati. Regarding to the similarity of the years 1991 and 1994 it is not possible to discuss the impact of the hydrological regime in this part of the Danube river.
Weevil taxocoenoses in the lower part have a similar course of abundance as the taxocoenoses in the middle part (Fig. 2, 3). The drop of the abundance in 1994 to the level in 1991 can be seen in the localities of GA and KU. The course of the abundance as well as the values of the average abundance are very similar in the areas of TO (middle part) and KU (lower part). These values indicate a similar character in habitat conditions (soil, humidity, nutritious plants) of the middle and the lower part of the monitoring territory.
The average abundance in all areas (soil of Salici - Populetum) shows a value of 19.2 ind.m-2, of which the highest value was observed in 1992. In the first and the last year of research similar average abundance values - 13.2 and 10.2 ind.m-2 , respectively (Fig. 4), were observed in all areas.
During the research I also observed changes in the number of species of weevil taxocoenoses. In all areas the average number of species was, in 1991 20, 26 in 1992, 21 in 1993, and 13 in 1994 (Table 3). The decrease in number of species in 1994 concerns mainly the geophilous species of the genus Dorytomus.
It is important to record the influence of exogenous environmental factors. The following factors observed were: pH value, contents of CaCO3, river floods and extreme summer droughts. Forest stand structure has only an influence on the composition of the following accompanying weevil taxocoenoses (Lignyodes sp., Stereonychus sp., Byctisus sp., Tanymecus sp., Curculio sp.).
The Danubian floods are a decisive phenomenon.The flood in the 1991 manifested itself most decisively. The abundance of weevils after the flood was low in each area (Fig. 1, 2, 3), because the soil humus layer was affected during the flood.
Only in the area KO the abundance exhibited a different course, because this area was not influenced by the floods. In 1992 a strong increase in the abundance occurred. The extreme drought in 1993 slightly decreased, or increased the abundance of weevils in the soil a little.
The year 1994, the second year after the starting of the supply of the river branches with water from the Dobrohost intake structure, is interesting. The abundance of weevils is in the areas KO, BO, TO, GA and KU lower by 50% in comparison to 1991. The highest abundance is, in the area KO, which is caused, however, by a single geobiont species, Barypeithes chevrolati.
Also represented there, are weevils preferring dry and wet habitats in the taxocoenoses. The species of the genus Trachyphloeus indicate dry, sandy soils. Their presence in the areas of KO, OL, and BO indicate a certain degree of a dry habitat.
On the contrary, the hygrophilous weevils Tanysphyrus, Bagous, Grypus, Rhinoncus, Phytobius, and Poophagus are represented in the taxocoenoses in the areas of DO, TO, GA, KU, and CI. There are good humid conditions in the soil in these areas. Geobionts (Trachyphleous, Brachysomus, Barypeithes, Acalles), which react to soil chemical composition, have an indication significance, that the soils are mostly calciphilous.
Some questions remain open:
Changes in weevil taxocoenoses are dependent upon a set of factors of exogenous as well as endogenous importance. For a relevant evaluation of the impact of the Hydroelectric power structure Gabcikovo on a part of the edaphon, a complex study using a precise quantitative method is necessary. Up until the present, results of the study of weevil taxocoenoses and their changes cannot be evaluated as an impact of the Hydroelectric power structure Gabcikovo. Only further studies will show more precisely the interpretation of the relation between the taxocoenose, changes and the impact.
| Species | Study area | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| KO | RU | OL | DO | BO | TO | GA | KU | CI | |
| Pselaphorhynchites tomentosus Gyll. | 1 | ||||||||
| Byctiscus betulae (L.) | 2 | ||||||||
| Apion frumentarium (L.) | 1 | ||||||||
| Apion urticarium (Hbst.) | 4 | 4 | 3 | 123 | 1 | 3 | |||
| Apion carduorum Kby. | 1 | 3 | 34 | ||||||
| Apion onopordi Kby. | 2 | 23 | 2 | ||||||
| Apion tenuae Kby. | 1 | 2 | 2 | ||||||
| Apion aestimatum Fst. | 1 | ||||||||
| Apion pisi (F.) | 2 | 12 | 2 | 2 | |||||
| Apion punctigerum (Payk.) | 2 | 4 | |||||||
| Apion virens Hbst. | 2 | ||||||||
| Apion assimile Kby. | 2 | ||||||||
| Apion nigritarse Kby. | 12 | 1 | 2 | ||||||
| Apion trifolii (L.) | 3 | ||||||||
| Otiorhynchus inflatus Gyll | 3 | ||||||||
| Otiorhynchus raucus (F.) | 1234 | 4 | 34 | 2 | 34 | ||||
| Otiorhynchus ovatus (L.) | 23 | 1 | 3 | ||||||
| Otiorhynchus velutinus Germ. | 3 | ||||||||
| Simo hirticornis (Hbst.) | 123 | 4 | 3 | ||||||
| Phyllobius sinuatus (F.) | 234 | 14 | 134 | 3 | |||||
| Phyllobius oblongus (L.) | 134 | 23 | 234 | 3 | |||||
| Phyllobius argentatus (L.) | 2 | ||||||||
| Phyllobius maculicomis Germ. | 2 | ||||||||
| Phyllobius pomaceus Gyll. | 3 | 3 | |||||||
| Phyllobius pyri (L.) | 2 | ||||||||
| Prachyphloeus scabriculus (L.) | 2 | ||||||||
| Trachyphloeus aristatus Gyll. | 12 | 4 | 1 | ||||||
| Trachyphloeus spinimanus Germ. | 3 | 4 | |||||||
| Trachyphloeus asperatus (Boh.) | 3 | ||||||||
| Trachyphloeus parallelus Seidl. | 123 | 3 | 3 | ||||||
| Polydrusus marginatus teph | 3 | ||||||||
| Polydrusus sericeus (Schall.) | 3 | 3 | 3 | ||||||
| Polydrusus pterygomalis Boh. | 3 | 4 | 2 | 3 | 4 | ||||
| Polydrusus corruscus Germ. | 13 | 1 | |||||||
| Sciaphilus asperatus (Bonsd.) | 3 | 3 | 134 | 3 | |||||
| Brachysomus echinatus (Bonsd.) | 1234 | 2 | 2 | 2 | |||||
| Barypeithes chevrolati (Boh.) | 1234 | 34 | 1234 | 2 | |||||
| Strophosoma melanogrammum Forst. | 2 | ||||||||
| Sitona inops Gyll. | 1 | ||||||||
| Sitona cylinricollis Fahrs. | 1 | ||||||||
| Sitona Iineelus (Bonsd.) | 1 | ||||||||
| Sitona suturalis Steph | 1 | ||||||||
| Sitona lineatus (L.) | 4 | 4 | 23 | 3 | |||||
| Sitona humeralis Steph. | 3 | 23 | 13 | ||||||
| Sitona hispidulus (F.) | 23 | 2 | 123 | 23 | 23 | ||||
| Sitona macularis (Mrsh.) | 134 | 34 | 1234 | 24 | 24 | 124 | 124 | 4 | |
| Tanymecus palliatus (F.) | 2 | ||||||||
| Tropiphorus carinatus (Mull;) | 12 | ||||||||
| Bothynoderes punctiventris (Germ) | 2 | ||||||||
| Dryophthorus corticalis (Payk.) | 3 | ||||||||
| Cossonus parallelopipedus (Hbst.) | 4 | ||||||||
| Cossonus linearis (F.) | 3 | 3 | 13 | 23 | 4 | ||||
| Phloephagus lignarius (Mrsh.) | 3 | ||||||||
| Stereocorynes truncorum (Germ) | 12 | ||||||||
| Hexathrum capitulum (Woll.) | 2 | ||||||||
| Bagous limosus (Gyll.) | 12 | 1 | |||||||
| Bagous subcarinatus (Gyll.) | 4 | 1 | 3 | ||||||
| Tanysphyrus lemnae (F.). | 1 | 12 | 123 | 12 | 3 | ||||
| Dorytomus longimanus (Forts.) | 234 | 3 | 1234 | 1234 | 23 | 23 | 1234 | 3 | |
| Dorytomus rufatus (Bed.) | 2 | 2 | |||||||
| Dorytomus melanophtahnus (Payk.) | 1 | 1 | 23 | 3 | 3 | ||||
| Dorytomus ictor (Hbst) | 124 | 34 | 123 | 1234 | 123 | 34 | 1234 | 34 | |
| Dorytomus tremulae (F.) | 4 | 3 | 1234 | 234 | 23 | 12 | 234 | 4 | |
| Dorytomus minutus (Gyll.) | 1234 | 3 | 123 | 3 | 24 | 3 | 2 | ||
| Dorytomus nordenskioeldi Fast. | 2 | 2 | 2 | 1 | 12 | ||||
| Dorytomus hirtipennis Bed. | 2 | 12 | |||||||
| Dorytomus schoenherri Faust | 2 | 23 | 2 | 3 | 2 | 24 | |||
| Dorytomus nebulosus (Gyll.) | 34 | 1234 | 1234 | 23 | 123 | 123 | 3 | ||
| Dorytomus villosulus (Gyll.) | 1 | ||||||||
| Dorytomus dejeani Fst. | 3 | 3 | |||||||
| Dorytomus affinis (Payk.) | 4 | 4 | 24 | 24 | 34 | 2 | 24 | ||
| Dorytomus rnajalis (Payk.) | 1 | ||||||||
| Dorytomus filirostris (Gyll.) | 3 | 234 | 1234 | 1234 | 1234 | ||||
| Dorytomus taeniatus (F.) | 2 | 3 | 123 | 23 | 3 | 2 | 23 | 3 | |
| Dorytomus puberulus (Boh.) | 1234 | 4 | 34 | 1234 | 123 | 234 | 1234 | 1234 | 34 |
| Grypus brunneirostris (F.) | 2 | ||||||||
| Ellescus scanius (Payk.) | 1 | ||||||||
| Ellescus scanius (L.) | 2 | ||||||||
| Lignyodes enucleator (Panz.) | 13 | 4 | 4 | ||||||
| Lignyodes muerlei Ferr. | 3 | ||||||||
| Lignyodes uniformis Desbr. | 1 | ||||||||
| Tychius cuprifer (Panz.) | 2 | ||||||||
| Tychius picirostris (F.) | 1 | ||||||||
| Anthonomus pedicularius (L.) | 23 | 3 | 1 | ||||||
| Anthonomus rubi (Hbst.) | 1 | 4 | 4 | ||||||
| Brabybatus creutzeri (Germ.) | 1 | ||||||||
| Curculio salicivorus Payk. | 1 | 2 | |||||||
| Curculio crux F. | 1 | ||||||||
| Trachodes hispidus (L.) | 123 | 1 | |||||||
| Lepyrus palustris (Scop.) | 1 | ||||||||
| Leiosoma detlexum (Panz.) | 2 | ||||||||
| Gronops inaequalis Boh. | 1 | ||||||||
| Sitophilus oryzae (L.) | 3 | ||||||||
| Acalles camelus (F.) | 1 | ||||||||
| Acalles hypocrita Boh. | 123 | 1 | 2 | ||||||
| Acalles echinatus (Germ.). | 1234 | 1234 | 1234 | 1 | 234 | 2 | 34 | ||
| Baris lepidü Germ. | 1 | ||||||||
| Phytobius comari (I-bst.) | 1 | ||||||||
| Rhinoncus incospectatus (Hbst.) | 1 | ||||||||
| Rhinoncus bruchoides (Hbst.) | 2 | ||||||||
| Rhinoncus pericarpius (L.) | 1 | ||||||||
| Rutidosoma globulus (Hbst.) | 4 | 4 | 4 | ||||||
| Amalorhynchus melanarius (Steph.) | 3 | 2 | 2 | 2 | |||||
| Poophagus sisymbrü (F.) | 1 | 1 | |||||||
| Ceutorhynchus sophiae (Stev.) | 1 | 1 | 2 | ||||||
| Ceutorhynchus floralis (Payk.) | 123 | 1234 | 1 | 234 | 1 | ||||
| Ceutorhynchus constrictus (Mrsh.) | 4 | 4 | |||||||
| Ceutorhynchus scrobicollis Ner,Wag. | 1 | 1 | 13 | 3 | 4 | 34 | |||
| Ceutorhynchus pallidactilus (Mrsh.) | 124 | 4 | 34 | 124 | 1234 | 2 | 123 | 124 | |
| Ceutorhynchus obstrictus (Mrsh) | 4 | ||||||||
| Ceutorhynchus erysirni (F.) | 1234 | 1234 | 13 | 234 | 124 | 2 | |||
| Ceutorhynchus pyrrhorhyncus Mrsh. | 2 | ||||||||
| Nedyus quadrimaculatus (L.) | 1234 | 134 | 12 | 3 | |||||
| Mecinus pyraster (Hbst.) | 3 | 3 | |||||||
| Stereonychus fraxini (Deg.) | 123 | 34 | 3 | ||||||
| Rhynchaenus rufitarsis Germ. | 1 | ||||||||
| Rhamphus ox acanthae Mrsh. | 4 | ||||||||
Table 2 Values of similarity index (Sörensen's index) of weevil taxocoenoses on 6 areas.
| KO | DO | BO | TO | GA | KU | |
|---|---|---|---|---|---|---|
| 0.4 | 0.5 | 0.4 | 0.5 | 0.4 | KO | |
| 0.6 | 0.5 | 0.6 | 0.5 | DO | ||
| 0.6 | 0.7 | 0.5 | BO | |||
| 0.6 | 0.6 | TO | ||||
| 0.6 | GA | |||||
| KU |
| Study area | 1991 | 1992 | 1993 | 1994 | ||||
|---|---|---|---|---|---|---|---|---|
| sp | A | sp | A | sp | A | sp | A | |
| KO | 28 | 31 | 32 | 22.5 | 31 | 22.5 | 12 | 15.1 |
| RU | 9 | 2.8 | ||||||
| OL | 26 | 15 | 15 | 5.7 | ||||
| DO | 26 | 27 | 30 | 29 | 23 | 30 | 14 | 42 |
| BO | 29 | 11 | 25 | 27 | 24 | 43 | 22 | 10.6 |
| TO | 12 | 2 | 22 | 7.4 | 19 | 7 | 8 | 2.4 |
| GA | 12 | 3 | 19 | 9.8 | 19 | 13.3 | 14 | 4.2 |
| KU | 14 | 5 | 30 | 22.4 | 14 | 6.3 | 14 | 4.2 |
| CI | 16 | 7.8 | 8 | 7.8 | ||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}