CONCLUSIONS The changes in representation of individual ecological groups of Carabidae in the within-dike area of the Danube during the period 1986-1994 show that the drainage process has started in this area, and resulted in initial stages of community succession tending to rise of ecologically less valuable heterogeneous ecotonal communities. This succession started as early before the diversion of the Danube main flow as a result of long-term continuous decrease of average ground water level, climatic fluctuations and various human activities in the past. In 1992-1994, after the Danube diversion it was considerably accelerated. The further development of the Carabidae communities has three alternatives characterised briefly as
The studied communities are further characterised by a low dominance of hygrophilous species preferring eutrophic swampy substrates like Agonum moestum, Pterostichus nigrita, Pterostichus anthracinus or Oodes helopioides. Almost no xenocoenous species occurred in the studied localities until 1989-1990. The total one year catch in ten traps counts approximately 1000 individuals. The species number in individual sampling plots moves from 30 to 35 species. In spite of the almost total replacement of the original natural willow-poplar stands by monocultures of various poplar cultivars the state of the Carabidaeae communities might be considered as natural. Their structure corresponded to the cyclic catastrophic climax state of the ecosystems in the within-dike area.
The xenocoenous species in floodplain forests in the Danube within-dike area may be represented by two groups of species. The first group includes mesohygrophilous forest species, which do not occur in extensive floodplains or which inhabit only the sporadically flooded forest of the group of geobiocoens Ulmi Fraxineta carpinea, growing on floodplain margins. The second group includes heliophilous species inhabiting fields, meadows, ruderals, etc. These species occur in natural forests of any type at most only as occasional migrants in a very limited number of individuals. The mesohygrophilous forest species did not and do not occur in the within-dike zone, neither their spreading into studied areas is probable in the near future because of negligible representation, isolation and large distance of suitable migration sources. The species of the second group, represented in this article illustratively by the two most abundant and expansive species Pseudophonus rufipes and Poecilus cupreus (Tab. 1), did not occur, up to 1991, in the studied area in an extent exceeding the normal state in other natural forest ecosystems. 1n 1991 and 1992 P. rufipes started to occur more frequently and in a slightly higher number of individuals. However, after putting the Gabcikovo hydroelectric power plant into operation, in 1993, the number of individuals and occurrence frequency increased markedly and in P. rufipes this trend continued in 1994 too. The penetration of these species with its up to date intensity can not still influence the Carabid communities in the floodplain forests considerably, but it is to be considered as one of the serious warning signals of the start of degradation processes in these ecosystems. In the strong damaged floodplain forests in other places the xenocenous species represent even 40% of Carabids [4]. Their penetration is stimulated by the earlier fall of leaves caused by the dry weather in July and August and a gradual decrease of humidity and ground water level, clear cutting, construction of forest roads and other human activities in the studied area.
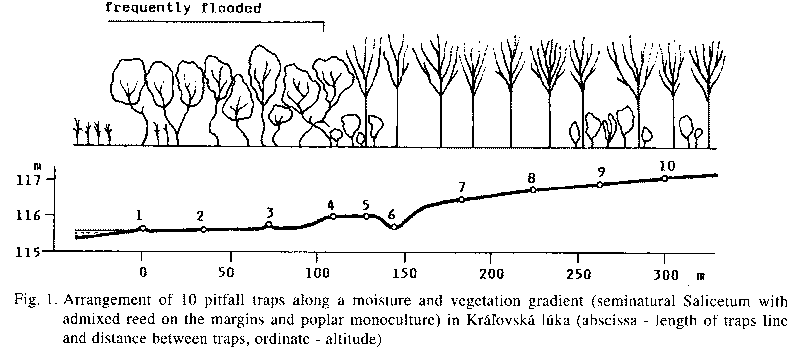
The decrease of abundance or local extinction of the most hygrophilous species represents a general tendency in the whole studied area. In this article it is illustrated by three strongly hygrophilous species of the genus Agonum (Tab. 3) in the sampling plot in Kralovska luka (Fig. 1). These species prefer permanently moist swampy habitats. The most hygrophilous A. fuliginosum occurred homogeneously along the whole ca. 400 m long sampling line in 1986-1989, when this locality was flooded more times a year for one or two weeks. Since 1990 this species disappeared at all. A little less hygrophilous Agonum micans left the moderately elevated part of the sampling plot and moved to the lower part around the remainder of a former oxbow (traps 1-4). The abundance of Agonum moestum, inhabiting in 1989 the moderately elevated part of the sampling line, and missing in the lower part being repeatedly flooded for a longer time, decreased already between 1986 and 1989, and similarly to the precedent species, this species moved to the moderately lower part around the oxbow shores. Bembidion dentellum is a characteristic inhabitant of the loamy shores of rivers. It is not a typical inhabitant of floodplain forests. It appeared here above after floods in 1987-88 and 1991-92. In 1989 it occurred homogeneously along the whole sampling line, while in 1992 and 1993 it concentrated in the lower parts.
The hygrophilous, but widely tolerant Carabus granulatus occurred, in the past, mostly in the moderately elevated area. Since 1990 it was spreading also to the moister lower parts. The moderately hygrophilous Pterostichus melanarius and P. niger occurred, until 1992, along the whole trapping line irregularly, and in a small number of individuals, with a slight preference for drier parts. After 1992, the abundance of both species increased strikingly along the whole sampling line, with an evident preference for the moister lower parts.
The high increase of the abundance of P. melanarius and P. niger in all study plots (Tab. 2) represents since 1991 a general trend. The increase of their abundance was made possible by more facts. Among the hygrophilous Carabids, P. niger and P. melanarius have the lowest requirements on moisture and both species are usually the first species, which indicate an increase of humidity in mesohygrophilous habitats. In addition, especially P. melanarius is an eurytopic tolerant species living frequently also in the fields and rudelars, and in a lower number of individuals, even in some more humid submontane forests. Both species are relatively good fliers and migrants, and in some cases they appear (or disappear) suddenly in high numbers without an apparent reason [5]. Furthermore, P. melanarius has, in contrast to almost all central European Carabids, a plastic reproductive cycle. All these properties allow them to colonise rapidly new habitats, even in the interior of the cities (Bratislava, Kamenne namestie square!![3]). It is probable that the rapid increase of their abundance in 1991 was stimulated also by the flood in August. Immediately after this flood, they belonged to the first species being active on extensive areas covered by deposited silt. Also immigrants from the adjacent fields might play a role.
It is to be accentuated that in the floodplain forest Horni les near Lednica na Morave P. niger, P. melanarius and Carabus granulatus were the only species, which tolerated the stop of regular spring floods and an average decrease of ground water level at 1.5 m [5].
Tab. 1. Increase in intensity of penetration of two xenocoenous Carabidae species into five floodplain forest communities in the within-dike area.
| Locality | Pseudophonus rufipes | Poecilus cupreus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1986 | 1989 | 1990 | 1991 | 1992 | 1993 | 1994 | 1986 | 1989 | 1990 | 1991 | 1992 | 1993 | 1994 | |
| Hrusov | - | 6 | 0 | 0 | 0 | 2 | 17 | - | 0 | 0 | 0 | 0 | 5 | 0 |
| Sulany | - | - | - | 1 | 6 | 8 | 7 | - | - | - | 0 | 3 | 0 | 0 |
| Bodicka brana | - | - | - | 1 | 0 | 6 | 7 | - | - | - | 0 | 0 | 1 | 1 |
| Kralovska luka | 1 | 0 | 0 | 1 | 0 | 0 | - | 0 | 2 | 2 | 0 | 0 | 17 | - |
| Istragov | - | 0 | 0 | 1 | 1 | 2 | - | - | 0 | 0 | 0 | 0 | 1 | - |
Tab. 2. Increase of the abundance of two moderately hygrophilous Carabidae species in five floodplain forest communities in the within-dike area.
| Locality | Pterostichus melanarius | Pterostichus niger | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1986 | 1989 | 1990 | 1991 | 1992 | 1993 | 1994 | 1986 | 1989 | 1990 | 1991 | 1992 | 1993 | 1994 | |
| Hrusov | - | 1 | 3 | 17 | 8 | 11 | 19 | - | 8 | 16 | 16 | 108 | 13 | 57 |
| Sulany | - | - | - | 172 | 46 | 501 | 117 | - | - | - | 31 | 194 | 487 | 86 |
| Bodicka brana | - | - | - | 140 | 208 | 553 | 261 | - | - | - | 140 | 592 | 390 | 201 |
| Kralovska luka | 16 | 7 | 6 | 28 | 71 | 233 | - | 10 | 55 | 11 | 13 | 147 | 368 | - |
| Istragov | - | 2 | 50 | 41 | 36 | 54 | - | - | 3 | 48 | 102 | 108 | 52 | - |
Tab. 3. Changes in the representation and spatial distribution of four strongly hygrophilous species (Agonum fuliginosum, A. micans, A. moestum and Bembidion dentellum) and of one tolerant (Carabus granulatus) and two moderately hygrophilous (Pterostichus niger and P. melanarius) species along the moisture and vegetation gradient in Kralovska luka.
| Species | Year | Sampling points on the moisture and vegetation gradient | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| A. fuliginosum | 1986 | 6 | 5 | - | 1 | 2 | - | 2 | 2 | 1 | - |
| 1989 | 1 | 2 | 5 | 1 | 7 | 2 | 1 | - | 1 | 2 | |
| 1990 | - | - | - | - | - | - | - | - | - | - | |
| 1991 | - | - | - | - | - | - | - | - | - | - | |
| 1992 | - | - | - | - | - | - | - | - | - | - | |
| 1993 | - | - | - | - | - | - | - | - | - | - | |
| A. micans | 1986 | - | - | - | - | 2 | 2 | - | - | - | - |
| 1989 | - | 1 | - | 8 | 10 | 8 | 3 | 3 | 1 | 1 | |
| 1990 | 4 | - | 2 | - | - | 5 | 2 | 1 | 2 | 1 | |
| 1991 | - | - | 2 | - | - | 6 | - | 1 | - | - | |
| 1992 | 9 | 5 | 6 | 12 | 2 | 5 | 1 | 1 | 1 | - | |
| 1993 | - | - | - | - | 9 | - | - | - | - | 2 | |
| A. moestum | 1986 | - | - | - | 7 | 12 | 38 | 8 | 3 | 5 | - |
| 1989 | - | - | - | - | 1 | - | 2 | 1 | 1 | - | |
| 1990 | - | - | - | 1 | 2 | 7 | 1 | - | 1 | - | |
| 1991 | - | - | - | - | 1 | - | - | 1 | 1 | - | |
| 1992 | 2 | 1 | - | - | 4 | - | 1 | - | 1 | - | |
| 1993 | - | 1 | 1 | - | 1 | - | - | - | - | - | |
| B. dentellum | 1986 | - | - | - | 1 | - | - | - | - | - | - |
| 1989 | 4 | 3 | 3 | 1 | 1 | 2 | 1 | 2 | 8 | - | |
| 1990 | - | - | - | - | - | - | - | - | - | - | |
| 1991 | - | - | - | - | - | 1 | 2 | 1 | - | - | |
| 1992 | 6 | 21 | 2 | 7 | - | - | - | 1 | - | - | |
| 1993 | - | 4 | 12 | 8 | 3 | 11 | - | - | - | - | |
| C.granulatus | 1986 | 7 | 1 | 3 | - | - | 21 | 8 | 2 | 1 | 1 |
| 1989 | - | 3 | 6 | 6 | 25 | 7 | 8 | 7 | 5 | 18 | |
| 1990 | 9 | 10 | 13 | 2 | 15 | 59 | 9 | 8 | 2 | 13 | |
| 1991 | - | 29 | 9 | 9 | 14 | - | 18 | 9 | - | - | |
| 1992 | 8 | 23 | 41 | 24 | 36 | 91 | 31 | 79 | 24 | 37 | |
| 1993 | 54 | 37 | 82 | 23 | 36 | 37 | 18 | 24 | 20 | 5 | |
| P. niger | 1986 | 1 | 1 | 2 | - | 2 | 9 | 1 | - | - | - |
| 1989 | - | - | - | - | 1 | - | 1 | 7 | 3 | 45 | |
| 1990 | - | - | 2 | 2 | - | 5 | 1 | - | - | - | |
| 1991 | - | - | - | - | - | 7 | 1 | 5 | - | - | |
| 1992 | 1 | 20 | 8 | 32 | 27 | 9 | 15 | 13 | 7 | 10 | |
| 1993 | 49 | 64 | 168 | 22 | 7 | 30 | 9 | 3 | 1 | 8 | |
| P. melanarius | 1986 | - | - | 1 | - | - | 15 | 1 | - | - | - |
| 1989 | - | - | - | - | 1 | - | - | 1 | - | 5 | |
| 1990 | - | - | - | - | - | - | - | 1 | 1 | 1 | |
| 1991 | 2 | - | 1 | - | 12 | 2 | 4 | 3 | 2 | 1 | |
| 1992 | - | 6 | 5 | 7 | 18 | 5 | 11 | 6 | 2 | 8 | |
| 1993 | 13 | 8 | 69 | 9 | 78 | 10 | 12 | 28 | 1 | 7 | |
{kind=link}